27/11/2025
Tóm tắt
Nghiên cứu này nhằm tổng hợp dữ liệu địa tầng và hóa thạch để phân tích quá trình phục hồi của nhóm Hai mảnh vỏ biển trong Trias ở miền Bắc Việt Nam. Các đặc điểm địa tầng chứa phức hệ Hai mảnh vỏ tiêu biểu, hình thái chức năng và hệ thống phân loại đã được phân tích, cho thấy tiến trình phục hồi Hai mảnh vỏ qua 3 giai đoạn của kỷ Trias theo hai xu hướng chính: phục hồi phân loại và phục hồi sinh thái. Các xu hướng này tương đồng với Nam Trung Quốc, tuy nhiên mức độ đa dạng ở miền Bắc Việt Nam thấp hơn do diện tích biển hẹp và quá trình phục hồi mạnh mẽ nhất chậm lại ở Trias giữa; từ Carni trở đi, điều kiện lục địa hóa hạn chế sự phát triển của nhóm hóa thạch này.
Từ khóa: Trias, Hai mảnh vỏ, phục hồi, cổ sinh thái, Bắc Việt Nam.
Ngày nhận bài: 2/10/2025; Ngày sửa chữa: 12/10/2025; Ngày duyệt đăng: 30/10/2025.
RECOVERY PATTERNS OF TRIASSIC MARINE BIVALVES IN NORTHERN VIETNAM
Abstract
This study aims to synthesize stratigraphic and fossil data to analyze the recovery of Triassic marine bivalves in northern Vietnam. The stratigraphic units containing characteristic bivalve assemblages, functional morphology, and taxonomic framework were analyzed, showing the recovery of bivalves through three stages of the Triassic according to two main parterns: taxonomic recovery and ecological recovery. These patterns are comparable with South China; however, the diversity level in northern Vietnam is lower due to the limited extent of marine basins, and the most pronounced recovery was delayed until the Middle Triassic. From the Carnian onward, continentalization restricted the development of fossil group.
Keywords: Triassic, bivalves, recovery, palaeoecology, Northern Vietnam.
JEL Classifications: O13, Q15, R00.
1. Mở đầu
Sự kiện tuyệt diệt Permi - Trias (khoảng 252 triệu năm trước) đã làm biến mất khoảng 80-95% sinh vật biển, gây ra sự sụp đổ toàn diện của các quần xã sống đáy và dẫn đến những thay đổi căn bản trong cấu trúc hệ sinh thái biển toàn cầu (Raup, 1979; Erwin, 1993; Benton & Twitchett, 2003). Sau thảm họa này, nhóm Hai mảnh vỏ - Bivalves (tổ tiên của các loài trai, hến, sò ngày nay) chiếm ưu thế, dần trở thành thành phần chi phối trong hệ sinh thái biển Mesozoi và đóng vai trò quan trọng trong tiến trình phục hồi sinh giới (Stanley, 1970; McRoberts, 2010).
Các nghiên cứu quốc tế cho thấy sự phục hồi của Hai mảnh vỏ diễn ra theo hai xu hướng chính: phục hồi phân loại (taxonomic recovery) và phục hồi sinh thái (ecological recovery). Phục hồi phân loại thường khởi phát sớm ngay trong Trias sớm (Indi-Olenek) với sự xuất hiện trở lại của một số giống “disaster taxa” như Claraia và Unionites, trong khi phục hồi sinh thái, tức là sự đa dạng hóa kiểu sống và chiến lược thích nghi, chỉ thực sự rõ nét từ Trias giữa, đặc biệt là Anisi-Ladin (Hautmann, 2006; Foster & Twitchett, 2014; Chen & Benton, 2012; Miao et al., 2025). Đến Trias muộn, các dạng sống chôn vùi và bán chôn vùi như Halobia và Bositra chiếm ưu thế, phản ánh rõ hơn cấu trúc đa dạng của hệ sinh thái biển (McRoberts, 2010).
Ở Việt Nam, khu vực Bắc bộ bảo tồn một loạt hệ tầng Trias biển với hóa thạch Hai mảnh vỏ phong phú (Hình 1). Tuy nhiên, so với Nam Trung Quốc, các xu hướng phục hồi Hai mảnh vỏ đã được nghiên cứu khá chi tiết, dữ liệu về tiến trình phục hồi ở miền Bắc Việt Nam còn phân tán và ít được tổng hợp một cách có hệ thống. Nghiên cứu này tập hợp và phân tích các dữ liệu địa tầng - cổ sinh ở Bắc Việt Nam nhằm làm rõ đặc điểm phục hồi của Hai mảnh vỏ trong Trias, tập trung vào ba giai đoạn: (1) Trias sớm; (2) Trias giữa - Carni và (3) Nori - Ret, góp phần làm sáng tỏ xu hướng phục hồi Hai mảnh vỏ biển trong kỷ Trias ở miền Bắc Việt Nam.
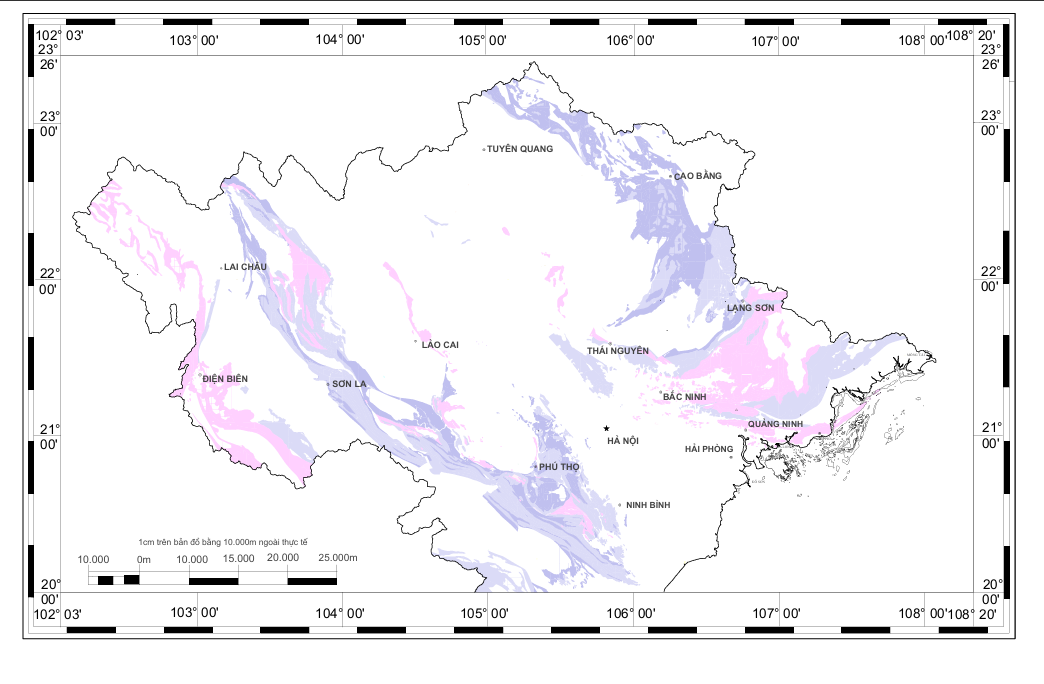
Hình 1. Sơ đồ phân bố trầm tích Trias ở Bắc Việt Nam (Trần Văn Trị và nnk., 2000)
2. Phương pháp nghiên cứu
Địa tầng và hóa thạch Hai mảnh vỏ Trias biển phân bố rải rác ở miền Bắc Việt Nam, bao gồm các trầm tích từ Trias sớm cho đến đầu Trias muộn ở bể Sông Hiến, bể An Châu và bể Sông Đà - Tú Lệ (Trần Văn Trị, 2009). Địa tầng và hóa thạch Trias biển từ lâu đã được nghiên cứu bởi các nhà địa chất kỳ cựu như Vũ Khúc, Trịnh Thọ, Đặng Trần Huyên, Nguyễn Đình Hữu, Nguyễn Kinh Quốc... Tất cả các nghiên cứu được tổng hợp trong công trình “Các phân vị địa tầng Việt Nam” trong đó đưa ra chi tiết về đặc điểm hệ tầng cũng như sự phân bố của hóa thạch Hai mảnh vỏ (Tống Duy Thanh, Vũ Khúc, 2011). Nghiên cứu tiến hành tóm tắt một số đặc điểm chính của địa tầng và hóa thạch Trias biển ở miền Bắc Việt Nam, cũng như bổ sung thêm một số nghiên cứu gần đây của Đặng Trần Huyên (2007); Komatsu (2006, 2007, 2008, 2010, 2017) và tài liệu hóa thạch của đề tài khoa học công nghệ "Nghiên cứu phân loại các loài của giống Costatoria, Trigonodus trong địa tầng Trias Trung ở một số mặt cắt ở Lạng Sơn (Đông Bắc bộ) và ý nghĩa địa tầng, cổ sinh thái và di sản địa chất của chúng", mã số: CS.2025.29. Đặc điểm hình thái chức năng được phân tích dựa trên các mô tả trong Atlas Cổ sinh vật Việt Nam, tập Thân mềm (Vũ Khúc và nnk., 2012) và Treatise on Inverterbrate Paleontology (Cox et. al., 1969). Đặc điểm hình thái thích nghi được phân tích dựa trên các nghiên cứu chuyên sâu về Hai mảnh vỏ của Stanley (1970, 1988). Luận giải phương thức sống của Hai mảnh vỏ dựa trên nghiên cứu về các kiểu sống của động vật đáy cổ sinh (sống trên bề mặt, chôn vùi nông, bán chôn vùi, chôn vùi sâu) và chiến lược dinh dưỡng liên quan của Brenchley & Harper (1998) và Dame, R. F. (2012).
3. Đặc điểm địa tầng và hóa thạch Hai mảnh vỏ Trias biển ở Bắc Việt Nam
3.1. Trias sớm
Trầm tích biển Trias sớm ở Bắc Việt Nam phân bố rải rác ở bể An Châu gồm hệ tầng Lạng Sơn với các trầm tích lục nguyên tuổi Indi và hệ tầng Bắc Thủy gồm các trầm tích lục nguyên, carbonat tuổi Olenek và bể Sông Hiến có các trầm tích phun trào hệ tầng Sông Hiến tuổi Indi - Olenek và trầm tích carbonat hệ tầng Hồng Ngài tuổi Olenek và hệ tầng Cò Nòi gồm các trầm tích lục nguyên chuyển tiếp lên carbonat ở bể Sông Đà (Trần Văn Trị và nnk. 2009, Tống Duy Thanh, Vũ Khúc, 2011). Hóa thạch Hai mảnh vỏ được ghi nhận khá phong phú, đặc biệt là các đới Claraia điển hình có ý nghĩa định tuổi địa tầng như đới Claraia stachei; đới Claraia wangi tuổi Olenek và đới Costatoria costata - Entolium discites microtis, đới Claraia aurita tuổi Indi (Vũ Khúc, 2012). Các loài của giống Claraia được ghi nhận bao gồm C. aff. wangi, C. aurita, C. kiparisova, C. vietnamica, C. stachei, C. griesbachi, C. yunnanensis, C. clarai, C. phobangensis, C. ex gr. stachei, C. gervilliaeformis, C. ex gr. aurita, C. desquamate, C. hubeiensis, C. intermedia, C. tridentina; C. aff. painkhadana. Đi cùng với Claraia thường có Eumorphotis (gồm E. multiformis, E. inaequicostata, E. venetiana, E. spinicosta, E. reticulate), Pteria (P. ussurica, Pteria sp.), Unionites (gồm U. canalensis, U. fassaensis), hiếm gặp hơn gồm Entolium, Posidonia, Velopecten, Gervillia, Hoernesia và Pecten (Đặng Trần Huyên, 1998, 2004, 2011).
3.2. Trias giữa - Carni
Trầm tích biển Asini - Carni ở Bắc Việt Nam lộ ra khá đầy đủ ở bể An Châu, lộ không đầy đủ ở các bể Quảng Ninh, Sông Hiến và Sông Đà - Tú Lệ (Trần Văn Trị, 2009). Giai đoạn Trias giữa, ở đới An Châu có các hoạt động tạo rift tuổi Anisi đã tạo nên hệ tầng Khôn Làng và trong thời Anisi muộn - Ladin các hệ tầng Điềm He và Nà Khuất được thành tạo trong môi trường lục nguyên, carbonat. Trong khi đó, các hệ tầng Yên Bình và Lân Páng với thành phần gồm các đá lục nguyên và carbonat được thành tạo ở các bể Sông Lô và Sông Hiến (Đặng Trần Huyên, 2008). Trong bể Sông Đà - Tú Lệ, trầm tích Anisi - Carni cũng được bảo tồn nhưng lộ ra không liên tục và bị phân cắt mạnh bởi các đới nâng kiến tạo. Bắt đầu với hệ tầng Đồng Giao (Anisi) gồm chủ yếu là trầm tích carbonat, chuyển tiếp lên trên là trầm tích lục nguyên hệ tầng Nậm Thẳm (Ladin); trầm tích phun trào hệ tầng Mường Trai tuổi Ladin muộn - Carni và cuối cùng là hệ tầng Nậm Mu và Pác Ma gồm các trầm tích lục nguyên tướng nước sâu tuổi Carni (Đặng Trần Huyên, 2011). Hai mảnh vỏ trong các hệ tầng giai đoạn này rất phong phú. Trong Anisi, phức hệ Costatoria - Trigonodus xuất hiện rộng rãi ở bể An Châu, , với các loài đặc trưng như Costatoria goldfussi, C. mansuyi, C. radiata, Trigonodus sandbergeri, T. tonkinensis, T. trapezoidalis, T. keuperinus, đi kèm Hoernesia inflata, Langsonella elongata, Pleuromya prosogyra, Pteria sturi. Bên cạnh đó, nhóm Daonella cũng bắt đầu phổ biến (D. laluensis, D. lindstroemi, D. elongata). Sang Ladin, phức hệ Costatoria - Trigonodus chiếm ưu thế, đi cùng có Daonella - Posidonia với các loài Daonella fluxa, D. lommeli, D. bulogensis, Posidonia wengensis, P. idriana. Đến Carni, các halobiid (Halobia comata, H. striatissima, H. superba, H. austriaca) và zittelihalobiid (Zittelihalobia superba, Z. rugosa) trở thành nhóm chủ đạo (Đặng Trần Huyên, 2007, 2008, 2011).
3.3. Trias muộn (Nori - Ret)
Trong giai đoạn Nori - Ret, các trầm tích biển lộ ra chủ yếu ở bể Quảng Ninh, đặc trưng bởi hệ tầng Hòn Gai; bể Sông Đà - Tú Lệ, đại diện bởi các hệ tầng Nậm Mu, Pác Ma và Suối Bàng. Đây là giai đoạn muộn nhất của biển Trias ở Việt Nam, bao gồm cả các trầm tích tướng biển sâu, lặng và thiếu oxy và các trầm tích gần bờ, đầm lầy chứa than. Hai mảnh vỏ biển sâu tiêu biểu gồm các dạng Halobia, Zittelihalobia, Burmesia; Hai mảnh vỏ biển nông được ghi nhận gồm: Unionites, Bakewellia, Gervillia, Hoernesia, Neoschizodus, Pleuromya, Modiolus, Palaeocardita, Songdaella, Langsonella, Costatoria, Trigonodus. (Vũ Khúc, 2011, 2012; Komatsu, 2017).
4. Hình thái thích nghi và kiểu sống của Hai mảnh vỏ Trias ở Bắc Việt Nam
Trong nghiên cứu cổ sinh thái, người ta sử dụng khái niệm hình thái chức năng để mô tả các đặc điểm hình thái của sinh vật phản ánh điều kiện và thói quen sống của sinh vật. Định nghĩa thói quen sống ở đây theo Stanley (1970) để thể hiện các hành vi của Hai mảnh vỏ liên quan đến đặc điểm bên ngoài với không gian sống, cách thức sống và sự tồn tại của chúng.
Có thể phân biệt ba nhóm kiểu sống chính. Sống trên bề mặt là những loài sống trên bề mặt nền đáy, chúng có thể gắn chặt vào vật cứng bằng sợi tơ chân bám trực tiếp bằng lớp vỏ hoặc chỉ nằm tự do trên bùn cát. Bán chôn vùi là các loài có phần thân vùi xuống trầm tích nhưng không hoàn toàn, thường sử dụng sợi tơ chân để bám vào các hạt trầm tích hoặc mảnh vụn trong nền đáy. Cuối cùng, chôn vùi là nhóm sống vùi trong trầm tích, có thể vùi nông với siphon ngắn hoặc vùi sâu với siphon dài; chúng thường thích nghi với hai chiến lược dinh dưỡng chính là lọc thức ăn lơ lửng trong cột nước hoặc khai thác chất hữu cơ trong trầm tích (Dame, 2012).
Trên cơ sở phân tích đặc điểm hình thái, ba kiểu sống của Hai mảnh vỏ kỷ Trias ở Bắc Việt Nam được tái dựng gồm dạng chôn vùi, bán chôn vùi và sống tự do trên bề mặt trầm tích (Hình 2).
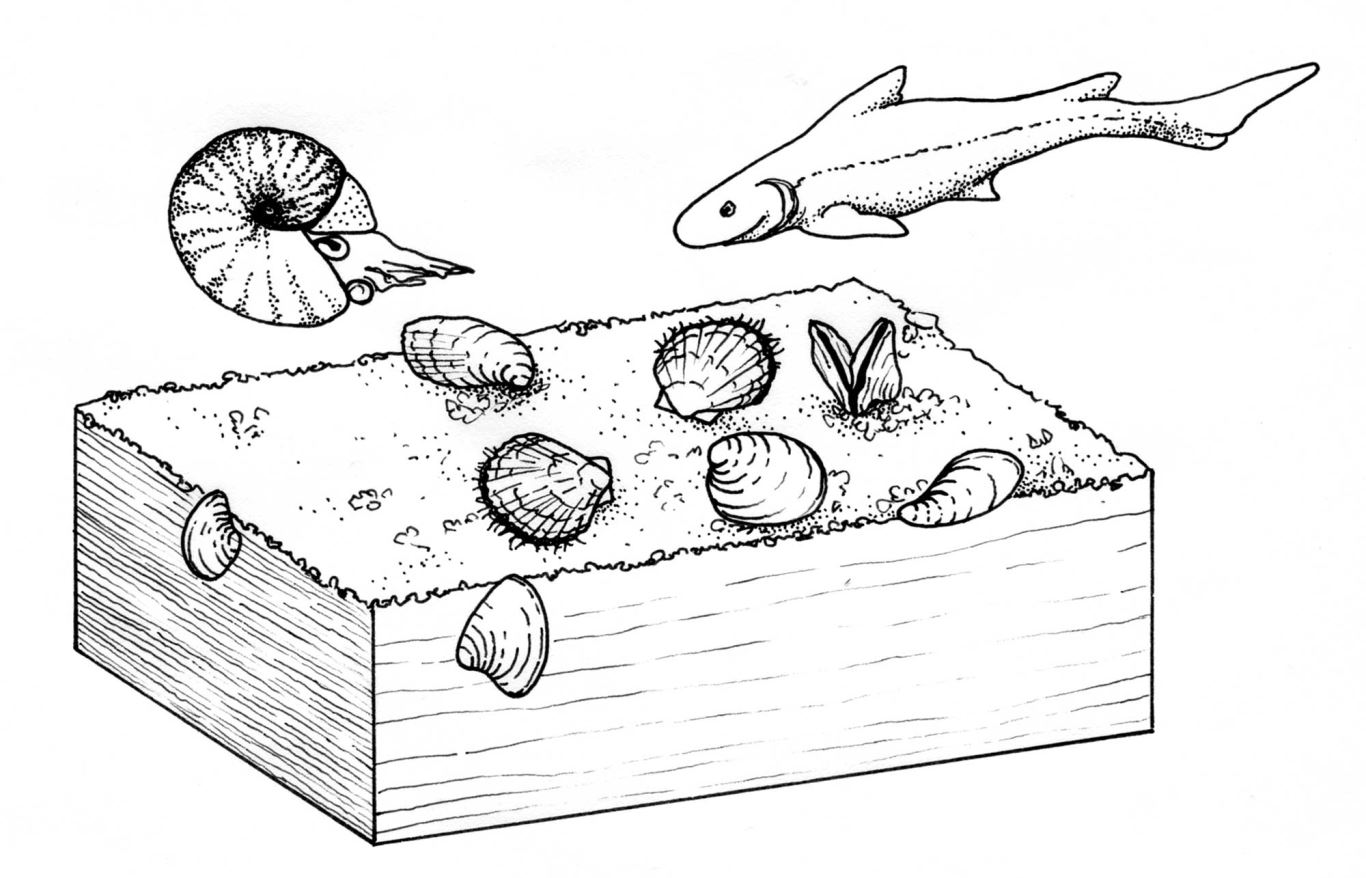
Hình 2. Các kiểu sống (life modes) của sinh vật đáy trong Trias sớm (theo Christi Sobel, 2020)
4.1. Trias sớm
Trong Trias sớm, sau sự kiện tuyệt diệt Permi - Trias, quần xã Hai mảnh vỏ ở Bắc Việt Nam được đặc trưng bởi các tập hợp nghèo nàn về phân loại nhưng có mật độ cá thể cao, chủ yếu do nhóm Claraia chiếm ưu thế tuyệt đối. Các loài thuộc nhóm này nhìn chung có vỏ nhỏ, mỏng, dẹt, hình bầu dục - tam giác, đỉnh thấp lệch trước, bề mặt nhẵn với vân tăng trưởng đồng tâm. Vỏ thường tạo tập hợp đơn loài, bảo tồn nguyên vẹn. Kiểu vỏ mỏng dẹt và đơn giản phản ánh chiến lược tiết kiệm năng lượng, thích nghi với điều kiện biển nông yếm khí, minh chứng cho vai trò nhóm loài bùng nổ số lượng sau sự kiện tuyệt diệt, thường có hình thái đơn giản, chiến lược sống phổ biến, chịu đựng tốt môi trường khắc nghiệt trong giai đoạn tồn tại sau sự kiện tuyệt chủng (Petsios & Bottjer, 2016). Với đặc điểm vỏ mỏng, dẹt, không có dấu hiệu cấu trúc bám hay khả năng đào vùi gợi ý Claraia có kiểu sống nằm tự do trên bề mặt bùn mềm (Yang et. al., 2001).
Sang Indi giữa và muộn, nhóm này vẫn duy trì ưu thế nhưng đã phát triển thêm nhiều loài mới như Claraia stachei, C. gervilliaeformis, C. wangi, C. griesbachi. Chúng vẫn duy trì kiểu sống tự do trên bề mặt bùn mềm nhưng đã tiến hóa thêm một số đặc điểm hình thái như mép vỏ thường có răng cưa hoặc gợn sóng nhẹ; bề mặt xuất hiện nếp gấp yếu xen với các vân tăng trưởng đồng tâm. Những đặc điểm này làm tăng độ bền cơ học, giúp chúng thích nghi với môi trường bùn - cát có động năng mạnh hơn so với giai đoạn Indi sớm (He et. al., 2013; Newell & Boyd, 1995). Nhóm Claraia chiếm ưu thế trong hệ sinh thái biển trong suốt thống Trias sớm.
|
|
|
Hình 3. Hóa thạch Claraia (trái) và Eumorphotis (phải) trong hệ tầng Hồng Ngài, vùng Hà Giang
Trong Onenek, một số loài của nhóm Eumorphotis cũng đã được ghi nhận, có thể kể đến như E. spinicosta, E. venetiana, E. inaequicostata và xuất hiện rải rác một số loài của nhóm Pteria và Unionites như Pteria ussurica và Unionites canalensis. Nhóm có vỏ mỏng, dẹt, có tai phát triển, đỉnh nhỏ lệch trước, những đặc điểm này phù hợp với kiểu sống bám vào nền cứng hoặc vỏ sinh vật khác. Ngược lại, Unionites có vỏ mảnh, đối xứng, ít gờ, phù hợp hơn với kiểu sống bán chôn vùi trong nền bùn mịn.
4.2. Trias giữa - Carni
Bước sang Trias giữa, trầm tích chứa Hai mảnh vỏ ở Bắc Việt Nam trở nên đa dạng từ biển ven bờ năng lượng cao, thềm lục địa nông cho đến ngoài khơi. Từ đó, số lượng giống loài và kiểu sống của Hai mảnh vỏ giai đoạn này cũng trở nên đa dạng và phong phú.
Trong các trầm tích biển nông ven bờ, đặc biệt chiếm ưu thế là nhóm Costatoria và Trigonodus. Nhóm Costatoria gồm những loài tiêu biểu như C. goldfussi, C. proharpa, C. pahangensis, C. vananensis. Chúng có đặc điểm chung là có vỏ chắc, gờ nan quạt thô, đỉnh nhô cao và mép vỏ dày, thích nghi với kiểu sống chôn nông trong nền bùn, cát và ổn định trong điều kiện năng lượng cao. Nhóm Trigonodus (T. sandbergeri, T. tonkinensis) thường có vỏ tam giác đến hình thoi, lồi mạnh, phần trước cắm xuống nền bùn, phần sau trồi lên, phản ánh kiểu sống bán chôn vùi, thích nghi tốt với môi trường sóng hoặc dòng chảy mạnh hơn (Vũ Khúc, 2012, Komatsu et al., 2010).
|
|
|
Hình 4 Hóa thạch Costatoria (trái) và Trigonodus (phải) trong hệ tầng Nà Khuất, vùng Lạng Sơn
Nhóm thứ hai chiếm ưu thế cao là Hoernesia, Langsonella, Asoella và Palaeolima, nhóm này gồm các dạng có vỏ trung bình, mép chắc và đôi khi có gân hướng tâm mảnh. Hoernesia có gờ nan quạt thô và vỏ chắc, thích nghi tốt với môi trường ven bờ năng lượng cao bằng kiểu sống chôn vùi nông. Langsonella (Hình 5), một giống đặc hữu của Bắc Việt Nam, có vỏ nhỏ, dẹt, nhẵn, được cho là sống trên bề mặt hoặc bán chôn vùi, thích nghi môi trường ít năng lượng ( Komatsu et al., 2010). Asoella có vỏ gần tam giác, gờ hướng tâm mảnh và đỉnh nhô, thích nghi bằng cách vùi nông và gắn nhẹ bằng tơ chân vào mảnh vụn sinh vật. Palaeolima, vỏ thuôn dài, gờ hướng tâm dày và vân đồng tâm, gắn bằng tơ chân vào nền cứng, thường tạo cụm tập trung (Komatsu et al., 2006; Aberhan, 1994).
Nhóm tiếp theo cũng thường xuất hiện là Pteria. Nhóm này gồm các loài có vỏ mỏng, dẹt, tai phát triển, đỉnh lệch trước, được cho là có kiểu sống gắn bằng tơ chân vào vật rắn hoặc vỏ sinh vật khác (Hình 5). Kiểu sống này đặc trưng cho các tướng ven bờ năng lượng vừa đến cao (Komatsu et al., 2006).
Nhóm cuối cùng chiếm ưu thế thấp hơn bao gồm Daonella, Posidonia, Entolium có vỏ mỏng, dẹt, đối xứng, các nghiên cứu trước đây cho rằng chúng có thể sống kiểu nằm tự do hoặc trôi nổi trong môi trường năng lượng thấp, thích nghi với môi trường biển sâu - yếm khí (Schatz, 2005; Komatsu et al., 2006).
|
|
|
Hình 5. Hóa thạch Langsonella (trái) và Pteria (phải) trong hệ tầng Nà Khuất vùng Lạng Sơn
Sang Carni, hai hệ sinh thái biển xuất hiện thêm các giống Halobia, Bositra và Palaeoneilo, đặc trưng cho tướng biển sâu yếm khí. Trong đó Halobia có vỏ mỏng, dẹt, trơn; Bositra vỏ nhỏ, mảnh, nhẵn; Palaeoneilo có vỏ hình bầu dục dày hơn, chúng thích nghi tốt bằng kiểu sống tự do trên bề mặt hoặc bán chôn vùi, thích nghi với đáy mềm, giàu vật chất hữu cơ (Komatsu et al., 2017; Del Piero, 2020; Ros Franch et al., 2014)
4.3. Trias muộn (Nori - Ret)
Nhóm Hai mảnh vỏ phổ biến nhất gồm các đại diện của họ Halobiidae và Posidoniidae, tiêu biểu là Halobia austriaca, H. talauna, H. cf. septentrionalis, H. superba, cùng với Zittelihalobia superba, Z. rugosa, Z. comata, Z. posterolaevis, Bositra sp… Các loài thuộc nhóm này có vỏ mỏng, dẹt, đối xứng, trơn nhẵn hoặc chỉ có vân tăng trưởng đồng tâm mảnh, gợi ý kiểu sống chiến lược sống tự do trên bề mặt hoặc chui rúc trong bùn sét giàu vật chất hữu cơ (Nakazawa & Nogami, 1967).
Ở tướng nước nông gần bờ, nhóm thích nghi môi trường ven bờ như Unionites damdunensis, U. convexa, Triaphorus gonoides, Palaeocardita globiformis, Neoschizodus songdaensis, Modiolus frugi và Bakewellia cf. magnissima chiếm ưu thế và một số loài kém ưu thế hơn có Palaeoneilo sp., Leda timorensis và Burmesia phongthoensis. Các loài trong tập hợp này có đặc điểm chung là vỏ dày hơn, hình bầu dục hoặc tam giác, mép vỏ lượn sóng, thích nghi với chôn vùi nông hoặc gắn nhẹ trong nền bùn - cát ven bờ.
5. Thảo luận
5.1. Đặc điểm và xu hướng phục hồi Hai mảnh vỏ Trias ở Bắc Việt Nam
Tiến trình phục hồi của Hai mảnh vỏ sau tuyệt diệt Permi - Trias (PTME) thể hiện rõ nét qua ba giai đoạn lớn, phản ánh cả quá trình phục hồi phân loại và phục hồi sinh thái.
Phục hồi phân loại: Trong giai đoạn Trias sớm (Indi - Olenek), thành phần Hai mảnh vỏ nghèo nàn, hầu như chỉ gồm Claraia (các loài C. aurita, C. vietnamica, C. concentrica) cùng một số loài phụ trợ như Unionites, Eumorphotis. Đây là giai đoạn sinh tồn sau PTME, phản ánh sự tồn tại của các loài bùng phát với đa dạng thấp (Komatsu, 2006, 2008). Bước sang Trias giữa (Anisi - Ladin), số lượng giống loài tăng mạnh, với sự xuất hiện của nhiều giống điển hình như Costatoria, Trigonodus, Hoernesia, Langsonella, Pteria, Asoella, Palaeolima. Giai đoạn này cho thấy sự bùng nổ đa dạng phân loại đạt đến cực đại, có ý nghĩa lớn trong phục hồi Hai mảnh vỏ tại miền Bắc Việt Nam. Đến Trias muộn (Nori - Ret), thành phần phân loại không bùng nổ về số lượng như Anisi, nhưng vẫn ghi nhận sự hiện diện ổn định của các loài mới (Halobia, Bositra, Palaeoneilo).
Phục hồi sinh thái: Trong Trias sớm, các tập hợp Claraia chỉ một kiểu sống duy nhất là sống trên bề mặt. Chúng vỏ mỏng, thích nghi với điều kiện môi trường bất ổn và oxy thấp (Komatsu et al., 2008). Sang đến Anisi - Ladin, Hai mảnh vỏ giai đoạn này xuất hiện nhiều kiểu thích nghi khác: Bán chôn vùi và chôn vùi nông (Costatoria, Trigonodus, Hoernesia, Langsonella); kiểu bám đáy (Pteria, Asoella); kiểu hỗn hợp - bám đáy và sống tự do trên bề mặt (Palaeolima). Sự đa dạng về kiểu sống chứng tỏ Hai mảnh vỏ đã đạt được phục hồi sinh thái rõ rệt. Sự phục hồi này tiếp tục giữ cân bằng cho đến Carni, khi chiếm ưu thế vẫn là các dạng có kiểu sống tự do trên mặt đáy như Halobia, Zittelihalobia; kiểu sống chôn vùi và chôn vùi nông như Burmesia và Palaeoneilo, kiểu sống bám đáy bằng tơ chân như Gervillia, Bakewellia. Đến Nori - Ret, biển bị thu hẹp dần vào do các khối Cimmerian di chuyển dần về phía bắc (Metcalfe, 2013), Hai mảnh vỏ biển sâu vẫn duy trì kiểu sống tự do trên bề mặt hoặc chui rúc xuống đáy bùn mềm trong khi Hai mảnh vỏ ở tướng nước nông phát triển hơn về kích thước và cấu trúc vỏ để chúng tồn tại tốt hơn trong điều kiện gần bờ động năng lớn.
5.2. Sự tương đồng của xu hướng phục hồi Hai mảnh vỏ ở Bắc Việt Nam và Nam Trung Quốc
Nhìn chung, các phức hệ hóa thạch Hai mảnh vỏ biển ở Trung Quốc và Bắc Việt Nam khá tương đồng (Bảng 1).
Tại Nam Trung Quốc, các nghiên cứu đã chỉ ra rằng phục hồi của Hai mảnh vỏ diễn ra theo hai pha rõ rệt. Trong Trias sớm (Indi - Olenek), quần xã rất nghèo nàn, gần như chỉ có Claraia và một số dạng đồng hành (Eumorphotis, Unionites) - nhóm “disaster taxa” điển hình, phản ánh giai đoạn sinh tồn trong điều kiện biển nông yếm khí (Hautmann, 2006; McRoberts, 2010; He et. al., 2007). Sang Anisi - Ladin, có sự bùng nổ về số lượng loài và chiến lược sinh thái: xuất hiện nhiều giống đặc trưng như Costatoria, Trigonodus, Daonella, Pteria, Asoella, đại diện cho các kiểu sống khác nhau (chôn vùi nông, bán chôn vùi, bám dính, trôi nổi). Đây là giai đoạn phát tỏa về mặt hình thái quan trọng, tái lập cấu trúc quần xã biển (Komatsu et al., 2010; Song & Dunhill, 2018). Đến Carni - Nori, nhóm Halobia, Zittelihalobia, Bositra chiếm ưu thế, phản ánh xu thế thích nghi với môi trường biển sâu hơn, nghèo oxy (McRoberts, 2010; Miao et al., 2025). Các nghiên cứu gần đây nhấn mạnh rằng ở Nam Trung Quốc, sự đa dạng phân loại và đa dạng sinh thái có xu hướng đồng bộ từ Anisi đến Carni, điều hiếm thấy ở quy mô toàn cầu (Foster & Twitchett, 2014; Miao et al., 2025).
|
Thống |
Bậc |
Bắc Việt Nam |
Trung Quốc |
|---|---|---|---|
|
Trias thượng |
Ret |
Cardinia - Unionites damdunensis |
Yunnanophorus - Trigondotus - Unionites |
|
Nori |
Burmesia - Halobia norica |
Burmesia lirata - Monotis salinaria; Halobia norica - H. distincta |
|
|
Carni |
Halobia talauana; Zittelihalobia superba |
Halobia pluriiradiata - H. convexa; H. rugosa - H. superba |
|
|
Trias trung |
Ladin |
Costatoria goldfussi - Trigonodus; Daonella indica |
Daonella bulogensis bifurcata - Mytilus eduliformis; D. variifurcata - D. indica |
|
Anisi |
Costatoria curvirostris; Daonella elongata |
Daonella moussou - Leptochondria illyrica; D. guizhouensis - Costatoria goldfussi |
|
|
Trias hạ |
Olenek |
Costatoria costata - Entolium discites microtis; Claraia aurita |
Pteria cf. murchisoni - Eumorphotis inaequicostata |
|
Indi |
Claraia stachei; Claraia wangi |
Eumorphotis multiformis; Claraia aurita, C. griesbachi, C. stachei, C. wangi, Pteria ussurica |
Bảng 1. Các đới hóa thạch Hai mảnh vỏ đặc trưng ở Bắc Việt Nam và Trung Quốc (theo Vũ Khúc, 2012 và Tong, 2018)
Ở miền Bắc Việt Nam, xu thế phục hồi có nhiều điểm tương đồng nhưng cũng có những khác biệt so với Nam Trung Quốc. Giai đoạn Indi cũng ghi nhận sự thống trị của Claraia aurita, C. stachei, C. wangi trong các tập hợp đơn loài, phản ánh chiến lược sống tự do trên bề mặt. Giai đoạn Anisi - Ladin, miền Bắc Việt Nam xuất hiện thêm một loạt các giống mới Costatoria, Trigonodus, Hoernesia, Langsonella, Daonella, Pteria, Asoella, với kiểu sống đa dạng, cho thấy quá trình mở rộng sinh thái mạnh mẽ tương tự Nam Trung Quốc (Komatsu et al., 2010). Tuy nhiên, từ Carni trở đi, trầm tích lục địa chiếm ưu thế ở Bắc Việt Nam, nên chỉ ghi nhận hạn chế các tập hợp Halobia - Bositra trong một số hệ tầng (Suối Bàng, Nậm Mu, Pác Ma), trước khi chuyển hẳn sang các giống Hai mảnh vỏ ven bờ với Unionites, Cardinia vào Nori - Ret.
|
Claraia wangi |
Claraia aurita |
Claraia stachei |
|
Entolium discites microtis |
Daonella indica |
Unionites damdunensis |
|
Halobia norica |
Halobia talauana |
Zittelihalobia superba |
Hình 6. Hình ảnh minh họa của một số hóa thạch Hai mảnh vỏ Trias (theo Komatsu, 2008 và Vũ Khúc, 2012)
So sánh hai khu vực cho thấy, ở Nam Trung Quốc, phục hồi cả về phân loại và sinh thái đạt mức hoàn chỉnh, kéo dài liên tục đến Trias muộn với sự thay thế tuần tự các phức hệ. Trong khi đó, ở miền Bắc Việt Nam, quá trình phục hồi mạnh mẽ nhất dừng lại ở Trias giữa; từ Carni trở đi, điều kiện lục địa hóa hạn chế sự phát triển của các cộng đồng biển sâu điển hình.
6. Kết luận
Quá trình phục hồi của Hai mảnh vỏ (Bivalvia) trong Trias biển ở miền Bắc Việt Nam phản ánh hai xu thế chính: (i) Phục hồi phân loại bắt đầu sớm từ Indi với các tập hợp đơn điệu của Claraia, đạt đỉnh ở Anisi - Ladin với sự đa dạng mạnh mẽ của Costatoria, Trigonodus, Hoernesia, Langsonella, Pteria và Asoella; (ii) Phục hồi sinh thái thể hiện rõ ở Trias giữa - Carni với sự xuất hiện của nhiều kiểu sống (trên bề mặt, bán chôn vùi và chôn vùi nông). Đến Nori - Ret, sự phục hồi sinh thái không phát triển thêm mà chỉ duy trì các phức hệ với lối sống tương đồng với Trias giữa.
So sánh với Nam Trung Quốc cho thấy xu thế phục hồi cơ bản tương đồng: Trias sớm nghèo nàn với Claraia, Trias giữa đa dạng hóa mạnh mẽ về phân loại và sinh thái, Trias muộn xuất hiện các nhóm Halobia - Bositra phản ánh xu thế chuyển đổi lối sống từ kiểu sống tự do trên bề mặt đáy đến bán chôn vùi và chôn vùi nông. Tuy nhiên, khác biệt nổi bật là ở Việt Nam, sự phục hồi mạnh mẽ nhất dừng lại ở cuối Trias giữa; từ Carni trở đi, điều kiện biển hẹp và lục địa hóa đã giới hạn sự phát triển của các phức hệ biển sâu điển hình.
Lời cảm ơn: Nghiên cứu được tài trợ bởi đề tài khoa học công nghệ "Nghiên cứu phân loại các loài của giống Costatoria, Trigonodus trong địa tầng Trias Trung ở một số mặt cắt ở Lạng Sơn (Đông Bắc bộ) và ý nghĩa địa tầng, cổ sinh thái và di sản địa chất của chúng", mã số: CS.2025.29.
Trịnh Thái Hà, Nguyễn Thị Hồng Nhung, Đinh Công Tiến, Nguyễn Đức Thái, Nguyễn Đức Phong
Viện Khoa học Địa chất và Khoáng sản
(Nguồn: Bài đăng trên Tạp chí Môi trường số Chuyên đề Khoa học - Công nghệ IV/2025)
Tài liệu tham khảo
1. Aberhan, M. (1994). Paleoecology of the Middle Triassic bivalve communities from the Cassian Formation of the Dolomites (Northern Italy). Palaeogeography, Palaeoclimatology, Palaeoecology, 111(1-2), 1-26.
2. Allmon, W. D., & Mikkelsen, P. M. (Eds.). (2020). Bivalvia (by Christi Sobel). In Digital Atlas of Ancient Life. Retrieved from https://www.digitalatlasofancientlife.org/learn/mollusca/bivalvia/.
3. Benton, M. J., & Twitchett, R. J. (2003). How to kill (almost) all life: The end-Permian extinction event. Trends in Ecology & Evolution, 18(7), 358-365.
4. Brenchley, P. J., & Harper, D. A. T. (1998). Palaeoecology: Ecosystems, environments and evolution. Springer US.
5. Chen, Z. Q., & Benton, M. J. (2012). The timing and pattern of biotic recovery following the end-Permian mass extinction. Nature Geoscience, 5(6), 375-383.
6. Cox, L. R., et al. (1969). In R. C. Moore (Ed.), Treatise on Invertebrate Paleontology, Part N. Mollusca 6. Bivalvia. Geological Society of America & University of Kansas Press.
7. Damborenea, S. E., & Pagani, M. A. (2019). Early Jurassic protobranch bivalves from Chubut, Argentina. Journal of Paleontology, 93(5), 925-944.
8. Dame, R. F. (2012). Ecology of marine bivalves: An ecosystem approach (2nd ed.). CRC Press, Taylor & Francis Group.
9. Đặng Trần Huyên (2004). Tài liệu mới về địa tầng và cổ sinh các trầm tích T1 ở Tây Bắc bộ. Tạp chí Địa chất, A/283, 1-9.
10. Đặng Trần Huyên (2007). Địa tầng trầm tích Phanerozoi ở Đông Bắc bộ. Viện Khoa học Địa chất và Khoáng sản, Hà Nội.
11. Dang, T. H. (1998). Lower Triassic bivalves from the Hồng Ngài Formation, Sông Hiến Structural Zone. Journal of Geology (Vietnam), B/11-12, 95-106.
12. Đặng Trần Huyên, Nguyễn Đình Hữu, Toshifumi Komatsu (2008). Đới hóa thạch và việc đối sánh địa tầng các trầm tích Trias giữa ở Đông Bắc bộ. Tạp chí Địa chất, A/308, 1-8.
13. Del Piero, N., Rigaud, S., Takahashi, S., et al. (2020). Unravelling the paleoecology of flat clams: New insights from an Upper Triassic halobiid bivalve. Global and Planetary Change, 190, 103195.
14. Erwin, D. H. (1993). The Great Paleozoic Crisis: Life and death in the Permian. Columbia University Press.
15. Fang, Z. J. (2003). Permian-Triassic bivalves: A review of their systematics, biostratigraphy and palaeoecology. Geobios, 36(3), 299-312.
16. Foster, W. J., & Twitchett, R. J. (2014). Functional diversity of marine ecosystems after the Late Permian mass extinction event. Nature Geoscience, 7(3), 233-238.
17. He, W., Zhang, K., Yang, T., & Wu, S. (2013). Morphological evolution of Claraia species from the Late Permian (Changhsingian) to the Early Triassic (Induan) and the response to the Permian-Triassic stressed environment. In R. Rocha, J. Pais, J. C. Kullberg, & S. Finney (Eds.), STRATI 2013: First International Congress on Stratigraphy (pp. 1035-1038). Springer Geology.
18. Komatsu, T., Huyen, D. T., & Jin-Hua, Ch. (2008). Lower Triassic bivalve assemblages after the end - Permian mass extinction in South China and North Vietnam. Paleontological Research, 12(2):119-128.
19. Komatsu, T., Huyen, D. T., & Huyen, P. D. (2010). Radiation of Middle Triassic bivalves: Assemblages characterized by infaunal and semi-infaunal burrowers in a storm-and wave-dominated shelf, An Chau Basin, North Vietnam. Palaeogeography, Palaeoclimatology, Palaeoecology, 289(1-4), 1-17.
20. McRoberts, C. A. (2001). Triassic bivalves and the initial marine Mesozoic revolution: A role for predators? Geology, 29(4), 359-362.
21. McRoberts, C. A. (2010). Biochronology of Triassic bivalves. In S. G. Lucas (Ed.), The Triassic Timescale (Geological Society, London, Special Publications, Vol. 334, pp. 201-219). Geological Society of London.
22. Metcalfe, I. (2013). Gondwana dispersion and Asian accretion: Tectonic and palaeogeographic evolution of eastern Tethys. Journal of Asian Earth Sciences, 66, 1-33.
23. Miao, X., Tong, J., Huang, Y., Zhang, S., Li, P., Cao, Y., Chu, D., & Kiessling, W. (2025). Taxonomic and ecologic transitions in Triassic marine bivalve communities. PeerJ, 13, e19237.
24. Nakazawa, K., & Nogami, Y. (1967). Problematic occurrence of the Upper Triassic fossils from the western hills of Kyoto. Memoirs of the Faculty of Science, Kyoto University, Series of Geology and Mineralogy, 34(1), 9-22.
25. Petsios, E., & Bottjer, D. J. (2016). Quantitative analysis of the ecological dominance of benthic disaster taxa in the aftermath of the end-Permian mass extinction. Paleobiology, 42(3), 380-393.
26. Raup, D. M. (1979). Size of the Permo-Triassic bottleneck and its evolutionary implications. Science, 206(4415), 217-218.
27. Ros-Franch, S., Márquez-Aliaga, A., & Damborenea, S. E. (2014). Comprehensive database on Induan (Lower Triassic) to Sinemurian (Lower Jurassic) marine bivalve genera and their paleobiogeographic record. Paleontological Contributions, 8, 1-219.
28. Schatz, W. (2005). Palaeoecology of the Triassic black shale bivalve Daonella: An example from the Northern Calcareous Alps (Austria). Palaeogeography, Palaeoclimatology, Palaeoecology, 216(3-4), 189-201.
29. Sepkoski, J. J. (1981). A factor analytic description of the Phanerozoic marine fossil record. Paleobiology, 7(1), 36-53.
30. Song, H., Wignall, P. B., & Dunhill, A. M. (2018). Decoupled taxonomic and ecological recoveries from the Permo-Triassic extinction. Science Advances, 4(10), eaat5091.
31. Stanley, S. M. (1970). Relation of shell form to life habits of the Bivalvia (Mollusca). Geological Society of America Memoir, 125, 1-296.
32. Stanley, S. M. (2009). Earth system history (3rd ed.). W. H. Freeman.
33. Tong-Dzuy Thanh, Vu Khuc (Eds.) (2011). Stratigraphic Units of Vietnam. Vietnam National University Publisher.
34. Trần Văn Trị (Chủ biên) (2000). Bản đồ Địa chất Việt Nam, tỷ lệ 1:1.000.000. Cục Địa chất và Khoáng sản Việt Nam.
35. Trần Văn Trị, Vũ Khúc (2009). Địa chất và Tài nguyên Việt Nam. Nxb Khoa học Tự nhiên và Công nghệ.
36. Vũ Khúc (Chủ biên) (2012). Atlas Cổ sinh vật Việt Nam - Tập Thân mềm. Nxb Tài nguyên - Môi trường và Bản đồ VIệt Nam.
37. Vũ Khúc, Trịnh Thọ (1975). Những tài liệu cổ sinh và địa tầng Trias thu thập được trong 10 năm gần đây (1963-1972) và ý nghĩa của chúng. Tuyển tập Công trình nghiên cứu về Địa tầng (pp. 185-243). Nxb Khoa học và Kỹ thuật.
38. Wignall, P. B., & Hallam, A. (1991). Paleoenvironmental and stratigraphic distribution of organic-rich facies in the Triassic System. Palaeogeography, Palaeoclimatology, Palaeoecology, 93(4), 207-231.
39. Yang, F., Peng, Y., & Gao, Y. (2001). Study on the Late Permian Claraia in South China. Science in China Series D: Earth Sciences, 44, 797-807.