11/10/2022
TÓM TẮT:
Trồng rừng từ lâu đã được công nhận là một giải pháp hiệu quả để giảm tốc độ gia tăng CO2 trong khí quyển. Ngoài ra, các thông tin về sinh khối rừng và nồng độ carbon đóng vai trò quan trọng trong quản lý bền vững tài nguyên rừng. Đa số các khu rừng trồng chỉ gồm một loài cây duy nhất, tuy nhiên, xu hướng trồng rừng có từ hai loài cây trở lên đang được mở rộng. Kết quả của nghiên cứu này cho thấy, ô tiêu chuẩn (OTC) Marteloscope lưu trữ 172,80 tấn/ha sinh khối cây trên mặt đất và 86,40 tấn/ha carbon, đồng thời có sự đa dạng loài với độ đồng đều tương đối cao. Mặc dù OTC Marteloscope được cơ quan quản lý xác nhận là một khu rừng trồng, thông tin từ việc phân tích các chỉ số mô tả phân bố không gian cho thấy các cây trong khu vực nghiên cứu có xu hướng phân bố ngẫu nhiên. Kết quả của phân tích độ nhạy mô hình tối ưu thể hiện rằng khả năng tích lũy sinh khối và hấp thụ carbon sẽ giảm khi sự đa dạng loài tăng.
Từ khóa: Marteloscope, cấu trúc rừng, phân tích độ nhạy, sinh khối rừng.
Nhận bài: Ngày 16/9/2022; Sửa chữa: Ngày 21/9/2022; Duyệt đăng: Ngày 26/9/2022
1. Đặt vấn đề
Sinh khối rừng là nơi lưu dữ các bon trong khí quyển tồn tại dưới dạng các sản phẩm của thực vật như lá, thân, cành, rễ, … đồng thời tham gia vào chu trình các bon tự nhiên giúp cố định CO2 trong khí quyển từ đó hạn chế được hiện tượng nóng lên toàn cầu và hiệu ứng nhà kính.
Có nhiều phương pháp khác nhau nhằm tính toán các chỉ số đa dạng, tập trung vào làm rõ mối quan hệ giữa đa dạng loài và sự vận động của hệ sinh thái, với chủ đề quan trọng nhất là tìm hiểu mối quan hệ giữa năng suất, sức khỏe hệ sinh thái và sự đa dạng loài [1]. Bên cạnh các nghiên cứu về mối quan hệ giữa năng suất và đa dạng loài thì một chủ đề khác ít được nghiên cứu hơn là sự tương quan giữa cấu trúc lâm phần và năng suất.. Đối với lâm phần hỗn giao, ngoài sự đa dạng về loại cây, nhiều khía cạnh về cấu trúc lâm phần hỗn loài cũng hoàn toàn khác biệt so với lâm phần độc canh, dẫn đến năng suất cao hơn [2].
Đa số rừng trồng ở Việt Nam là rừng đơn loài cây duy nhất, nhưng xu hướng hiện nay là trồng rừng sử dụng hai hoặc nhiều loài cây ở giai đoạn trồng rừng để tận dụng không gian, ánh sáng,... Tại Việt Nam, các nghiên cứu đánh giá mối quan hệ đa dạng loài – năng suất rừng còn nhiều hạn chế. Bài viết đóng góp nghiên cứu về mối tương quan giữa sự đa dạng loài và cấu trúc lâm phần với sản lượng rừng tại huyện Thạch Thất, Hà Nội.
2. Dữ liệu và phương pháp
2.1 Thu thập dự liệu
Nhóm nghiên cứu đã thiết lập 1 ô tiêu chuẩn Marteloscope (OTC) với diện tích 1 hecta tại tọa độ 105° 30' 59.7852'' E, 20° 59' 47.9076'' N, trong khuôn viên Đại học Quốc gia Hà Nội tại Hòa Lạc, huyện Thạch Thất, Hà Nội. OTC được chia thành lưới gồm 16 ô nghiên cứu (ONC) (Hình 1).
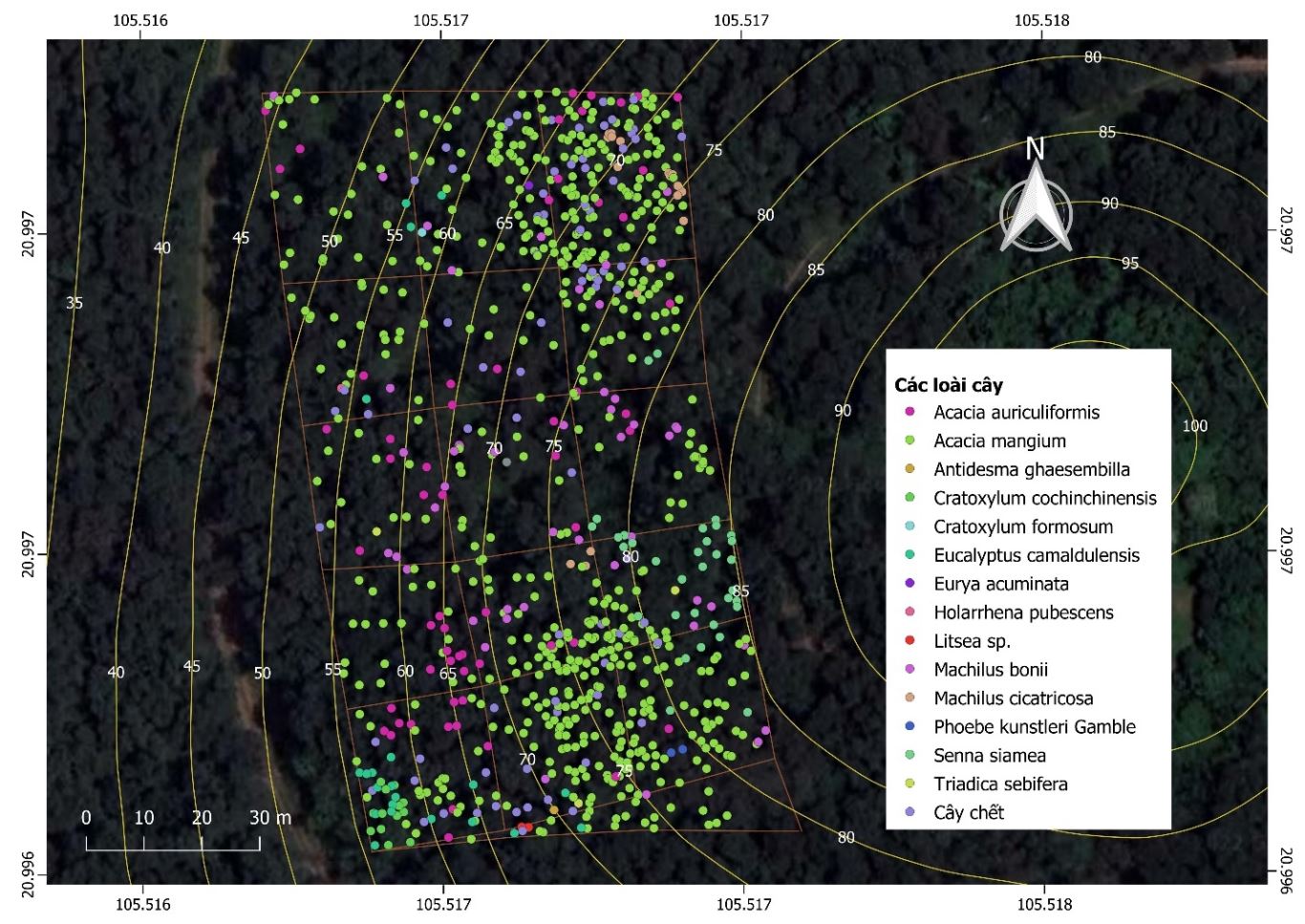
Hình 1. Bản đồ các loài cây tại OTC
Trong mỗi ONC, tọa độ cây, loài cây, chiều cao vút ngọn (H) và đường kính ngang ngực (DBH tại 1.3 m so với mặt đất) được xác định cho tất cả các cây gỗ có DBH > 5 cm. Sinh khối của cây được tính từ DBH và H, sử dụng các phương trình sinh khối thể hiện trong Bảng 1. Tỷ lệ phần trăm hàm lượng carbon (C) trong sinh khối khô được coi là 50% [3]. Vì vậy lượng carbon được tích lũy trong mỗi cây được tính như sau:
C (kg) = sinh khối × 0,5
Bảng 1. Phương trình sinh khối cho các loài khác nhau trong OTC

Các chỉ số đa dạng và chỉ số cấu trúc không được tính toán cho từng cây cá lẻ để ước tính mức độ phong phú, đa dạng của loài và phân tích định lượng toàn diện cấu trúc không gian của các loài cây trong khu vực nghiên cứu.
Chỉ số đa dạng Simpson (Simpson Biodiversity Index) thường được thể hiện là 1 – D, chỉ số này không chỉ đo về mức độ đa dạng mà xem xét cả độ giàu có và đồng đều với công thức tính:
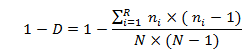
Trong đó: ni là số lượng cá thể của loài thứ i; N là tổng số cá thể của R loài. Giá trị gần tới 1 thì sự đa dạng cao và giá trị thấp gần 0 thì sự đa dạng thấp.
Chỉ số Shannon (Sh) tính đến cả sự phong phú và sự cân bằng của các loài hiện diện trong quần xã theo công thức:
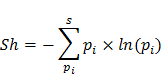
Trong đó: S là số loài, 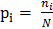 ; là số cá thể của loài i; N là tổng số cá thể của các loài trong lâm phần.
; là số cá thể của loài i; N là tổng số cá thể của các loài trong lâm phần.
Chỉ số ưu thế Berker-Parker (D) được xác định dựa vào tỷ lệ độ phong phú cao nhất theo công thức:
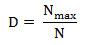
Trong đó: Nmax là số lượng cá thể của các loài phong phú nhất; N là tổng số cá thể trong lâm phần.
Chỉ số đồng đều (E) là chỉ số so sánh giá trị đa dạng thực tế với giá trị phân bố lớn nhất có thể theo công thức tính:
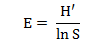
Trong đó: H′ là chỉ số Shannon; với là giá trị tối đa có thể của H′. Chỉ số đồng đều có giá trị từ 0 đến 1. Nếu chỉ số đồng đều càng gần 1 thì tất cả các loài chiếm ưu thế như nhau trong phân bố.
Độ hỗn loài (Mi) phản ánh mức độ tương đồng về loài giữa cây mục tiêu và các cây láng giềng gần nhất theo công thức:
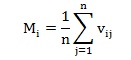
Trong đó: j là cây láng giếng so với cây mục tiêu i; nếu cây láng giềng và cây mục tiêu không cùng loài, ngược lại .
Bảng 2: Giá trị cụ thể, ý nghĩa sinh học của chỉ số
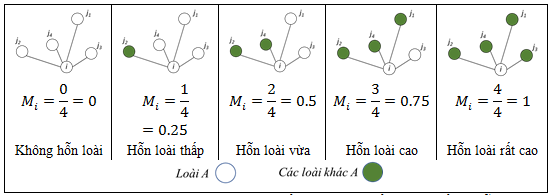
Tình trạng đa dạng không gian (MS) là chỉ số được nâng cấp từ chỉ số độ hỗn loài và không chỉ bao gồm sự pha trộn không gian mà còn tính toán cả số lượng các loài cây. MS được tính bằng công thức:
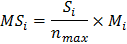
Trong đó: Si là số lượng các loài cây trong khu vực không gian của cây mục tiêu i, bao gồm cả cây i. nmax là số lượng loài tối đa trong đơn vị không gian này; là độ hỗn loài. . Một cây mục tiêu của một loài phổ biến có nhiều khả năng có cây lân cận cùng một loài nếu giá trị MS thấp và ngược lại.
Chi số phân biệt chiều cao (TH) mô tả sự khác biệt về chiều cao giữa cây tham chiếu i và n cây lân cận gần nhất j (j = 1 ... n). Chỉ số phân biệt chiều cao được tính như sau:
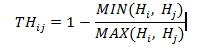
Trong đó: Hi, Hj là chiều cao của cây tham chiếu i và cây lân cận j; nếu TH = 1, cây i có sự khác biệt lớn về chiều cao so với cây j, nếu TH = 0 thì cây i và j có chiều cao bằng nhau.
Hàm Ripley L được sử dụng để phân tích mô hình phân bố không gian của các loài. Đối với n cây thử nghiệm với kích thước A, hàm L được tính như sau:
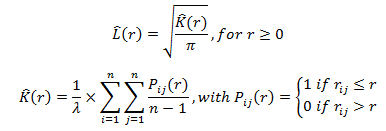
Trong đó: r là bán kính tập trung tại điểm điển hình (cây) i của mẫu điểm; rij là khoảng cách giữa cây i và cây j; λ = n/A đại diện cho mật độ điểm trung bình. Các dạng phân bố điểm có thể theo xu hướng hoàn toàn ngẫu nhiên (L̂(r) = 0), hoặc phân bố theo cụm (L̂(r) > 0) hoặc phân bố đồng đều (L̂(r) < 0).
Chỉ số tổng hợp (R) là tỷ lệ của khoảng cách cây lân cận gần nhất quan sát được trong mô hình với khoảng cách trung bình dự kiến trong phân bố cây ngẫu nhiên.
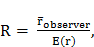
Trong đó: r̅ observer có nghĩa là khoảng cách giữa cây tham chiếu và cây lân cận; E(r) là khoảng cách trung bình được dự kiến khi cây được phân bố ngẫu nhiên; N là số của cây trong khu vực nghiên cứu; A là kích thước của khu vực nghiên cứu.
Chỉ số tổng hợp (R) nằm trong khoảng từ 0 (phân bố co cụm lớn nhất, tất cả các đối tượng phân bố tại cùng một điểm) và 2,1491 (phân phối đều).
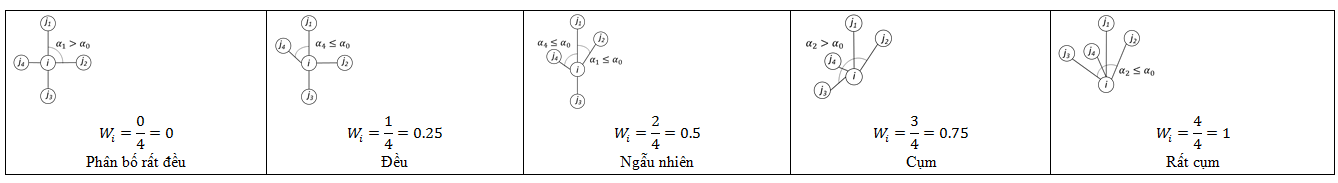
Chỉ số góc đồng nhất (W) mô tả mức độ đồng đều trong phân bố không gian của các cây riêng lẻ, dựa trên việc phân loại các góc giữa cây lân cận gần nhất của cây tham chiếu.
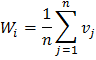
Trong đó: n là số lượng cây lân cận gần nhất xung quanh cây tham chiếu i; α0 = 360o /n là góc tiêu chuẩn dự kiến giữa cây tham chiếu và cây lân cận.
Bảng 3. Giá trị cụ thể ý nghĩa sinh học của chỉ số W
2.2. Phân tích thống kê
Phân tích thống kê được thực hiện đối với loài Keo tai tượng để có được cái nhìn sâu sắc hơn về mối quan hệ giữa độ phức tạp, đa dạng của lâm phần và sinh khối cây riêng lẻ. Các loài khác không được đưa vào quy trình phân tích thống kê do có kích thước mẫu quá nhỏ. Ba dạng phương trình tổng quan (PT.1 đến 3) đã được sử dụng để phát triển các mô hình khác nhau cho loài keo tai tượng, trong đó đại diện cho các tham số ước lượng và đại diện cho các biến giải thích khác nhau.
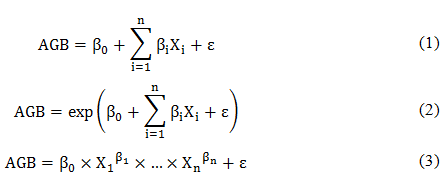
Phân tích tương quan cũng được thực hiện để kiểm tra đánh giá mối liên hệ tuyến tính giữa hai biến độc lập. Chỉ các cặp biến có hệ số tương quan (r) mới được đưa vào xây dựng các mô hình để tránh tính cộng tuyến. Việc lựa chọn phương trình tối ưu dựa trên tiêu chí thông tin Akaike (AIC), tiêu chí thông tin Bayes (BIC), hệ số xác định (R2) và sai số toàn phương trung bình (MSE). Các phân tích thống kê trong nghiên cứu này đều được thực hiện bằng phần mềm mã nguồn mở R phiên bản 3.5.3.
Sau khi lựa chọn được mô hình tốt nhất, nghiên cứu tiến hành phân tích độ nhạy của mô hình để đánh giá mức độ nhạy cảm của sinh khối cây trên mặt đất đối với sự biến động của các thông số và dữ liệu mà mô hình được xây dựng.
3. Kết quả nghiên cứu
3.1. Một số đặc trưng khu vực nghiên cứu
14 loài khác nhau đã được xác định trong khu vực nghiên cứu được thể hiện trong bảng 4 (Bảng 4).
Bảng 4. Một số đặc trưng các loài trong OTC
|
Danh pháp khoa học |
n |
DBH |
H |
AGB |
C |
CO2 |
||||||
|
Mean |
Median |
Max |
Min |
Mean |
Median |
Max |
Min |
|||||
|
Acacia mangium |
586 |
14.87 |
10.5 |
62.7 |
5 |
13.37 |
13.3 |
28.6 |
3.8 |
85.15 |
422.57 |
156.07 |
|
Acacia auriculiformis |
61 |
21.1 |
19.5 |
54.1 |
3.7 |
15.4 |
15.2 |
24 |
2.7 |
77.52 |
38.76 |
142.09 |
|
Cratoxylum cochinchinensis |
13 |
7.27 |
6.4 |
17 |
5.1 |
9.8 |
9 |
18 |
7 |
0.36 |
0.18 |
0.65 |
|
Eucalyptus camaldulensis |
21 |
11.48 |
12 |
25 |
5 |
12.92 |
13.8 |
19.6 |
7.3 |
1.14 |
0.57 |
2.09 |
|
Litsea sp. |
2 |
6.5 |
6.5 |
7 |
6 |
6 |
6 |
6.3 |
5.7 |
0.02 |
0.01 |
0.04 |
|
Machilus bonii |
47 |
8.48 |
7 |
57.4 |
5 |
6.7 |
6 |
24.6 |
3.3 |
3.41 |
1.71 |
6.25 |
|
Machilus cicatricosa S. Lee |
17 |
9.9 |
8.4 |
20.2 |
1.9 |
10.04 |
9.6 |
20 |
5.2 |
0.80 |
0.40 |
1.47 |
|
Phoebe kunstleri Gamble |
2 |
8.35 |
8.35 |
8.8 |
7.9 |
6.95 |
6.95 |
7.6 |
6.3 |
0.04 |
0.02 |
0.07 |
|
Senna siamea |
33 |
13.77 |
12.5 |
30.5 |
5.5 |
12.72 |
12.6 |
22.2 |
4.9 |
4.03 |
2.01 |
7.39 |
|
Triadica sebifera |
4 |
9.45 |
7.8 |
16.5 |
5.7 |
11.22 |
12.1 |
13.2 |
7.5 |
0.18 |
0.09 |
0.33 |
|
Antidesma ghaesembilla |
1 |
5 |
- |
- |
- |
3.1 |
- |
- |
- |
0.00 |
0.00 |
0.01 |
|
Cratoxylum formosum |
1 |
7.6 |
- |
- |
- |
5.1 |
- |
- |
- |
0.01 |
0.01 |
0.02 |
|
Eurya acuminata |
1 |
7 |
- |
- |
- |
10.1 |
- |
- |
- |
0.02 |
0.01 |
0.03 |
|
Holarrhena pubescens |
1 |
6.5 |
- |
- |
- |
5.8 |
- |
- |
- |
0.01 |
0.00 |
0.02 |
|
Cây chết |
79 |
10.23 |
8 |
35 |
4 |
8.58 |
8.75 |
18.5 |
1.9 |
- |
- |
- |
|
n: Tổng số cây mỗi loài; DBH: Đường kính ngang ngực (cm); Ht: Chiều cao vút ngọn (m); AGB: Tổng sinh khối cây trên mặt đất (tấn); C: Khối lượng các bon (tấn); CO2: Khối lượng carbon dioxide (tấn) |
||||||||||||
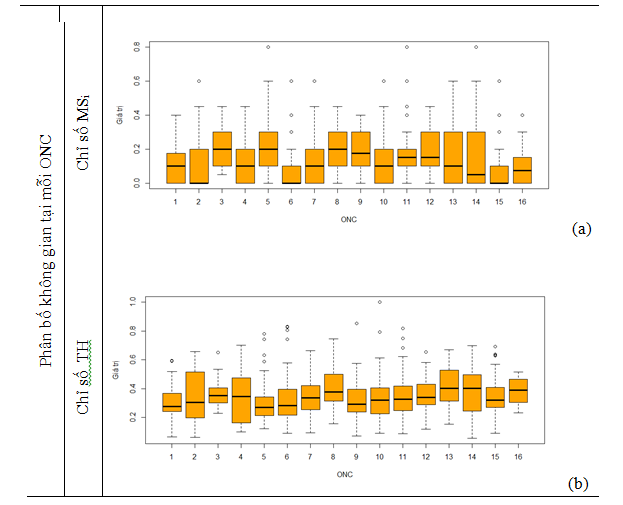
Kết quả tính toán mức độ phong phú và đa dạng của loài (hình 2), độ hỗn loài (hình 4a), phân bố cây theo chiều thẳng đứng (hình 4b) và chiều ngang (hình 5) cho thấy khu vực nghiên cứu có tính đa dạng loài cao với độ đồng đều tương đối cao. Mặc dù khu vực nghiên cứu được đơn vị quản lý xác nhận là rưng trồng nhưng các kết quả phân tích chỉ số phân bố không gian ngang (Hình 5) cho thấy rằng các cá thể trong OTC Marteloscope phân bố theo xu hướng ngẫu nhiên.
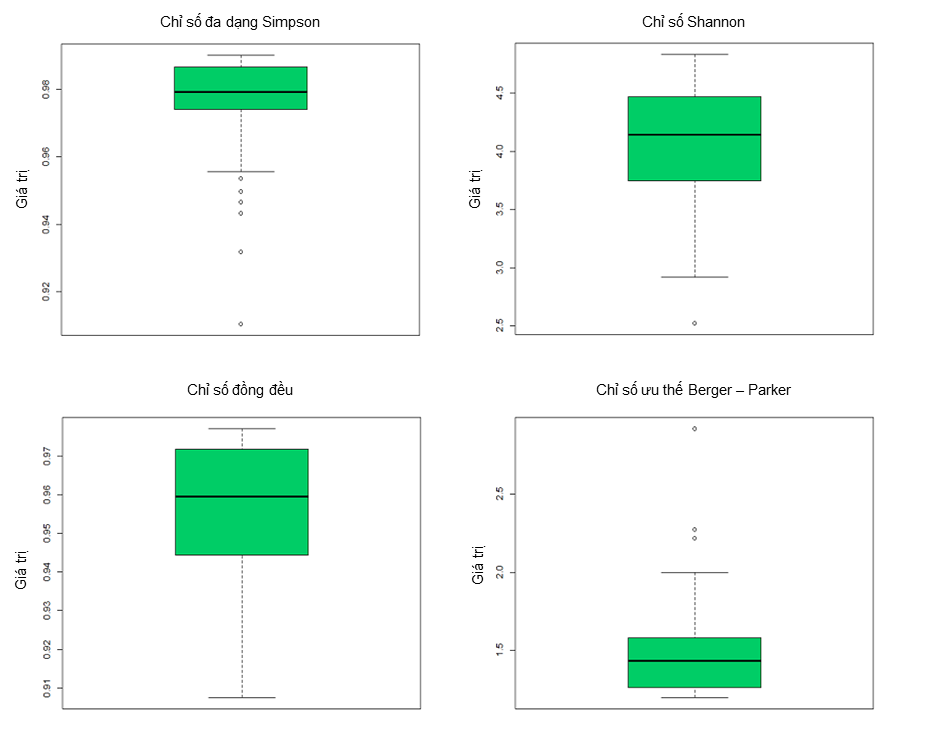
Hình 2. Các chỉ số đa dạng loài của OTC Marteloscope
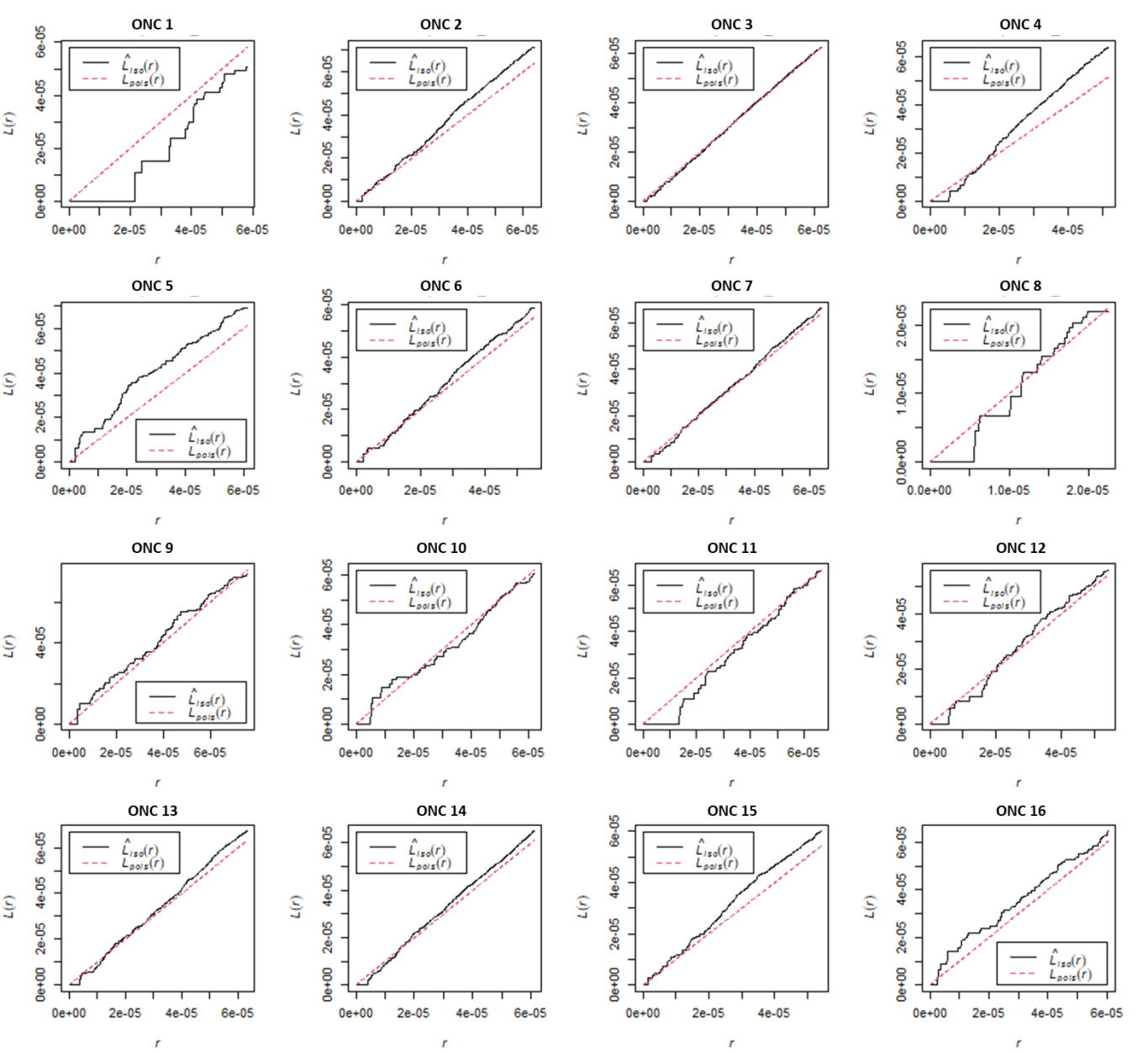
Hình 3. Kết quả phân tích hàm L cho từng ONC
3.2. Phân tích thống kê
Các mô hình khác nhau đã được phân tích và thử nghiệm, nhưng chỉ những mô hình đáp ứng các giả định hồi quy như tính đồng nhất của phương sai, tính chuẩn, tính tuyến tính và không tự tương quan với mức độ phù hợp cao mới được giữ lại trong nhóm mô hình khả dụng (Bảng 5).
Bảng 5. Cấu trúc 10 mô hình tốt nhất
|
STT |
Model |
|
|
|
|
|
p-value |
MSE |
AIC |
BIC |
|
1 |
|
1.377 |
1.684 |
9.934 |
- |
0.109 |
2.36E-15 |
1.607 |
1946.268 |
1963.693 |
|
2 |
|
5.377 |
1.747 |
-0.173 |
- |
0.109 |
2.40E-15 |
1.607 |
1946.298 |
1963.723 |
|
3 |
|
6.685 |
1.678 |
-0.690 |
- |
0.107 |
3.45E-15 |
1.609 |
1947.029 |
1964.453 |
|
4 |
|
15.827 |
1.691 |
-12.564 |
- |
0.106 |
5.44E-15 |
1.612 |
1947.943 |
1965.367 |
|
5 |
|
27.448 |
1.693 |
-24.173 |
- |
0.106 |
5.77E-15 |
1.612 |
1948.064 |
1965.488 |
|
6 |
|
5.345 |
1.68 |
-0.086 |
- |
0.105 |
7.15E-15 |
1.613 |
1948.494 |
1965.918 |
|
7 |
|
4.744 |
-0.009 |
0.340 |
- |
0.094 |
2.55E-13 |
1.633 |
1955.681 |
1973.105 |
|
8 |
|
4.240 |
1.870 |
-4.61E-05 |
- |
0.084 |
6.51E-12 |
1.651 |
1962.192 |
1979.61 |
|
9 |
|
2.571 |
-0.0006 |
1.795 |
1.660 |
0.060 |
5.89E-08 |
1.697 |
1979.293 |
2001.056 |
|
10 |
|
8.397 |
-0.0006 |
-4.011 |
1.664 |
0.060 |
6.57E-08 |
1.698 |
1979.521 |
2001.284 |
|
AGB sinh khối cây trên mặt đất (kg); TH: Chỉ số phân biệt chiều cao; Sh: chỉ số Shannon; Sm: chỉ số đa dạng Simpson; N: số cây/ha; E: chỉ số đồng đều; Mi: độ đồng đều; R: chỉ số tổng hợp. |
||||||||||
Đối với tất cả các mô hình, hệ số xác định (R2) ở mức thấp, cho thấy sự tương quan yếu giữa sinh khối trên mặt đất và các biến dự báo. Điều này là chấp nhận được bởi các mô hình này được xây dựng nhằm giải thích chứ không phải để dự đoán mối quan hệ giữa sinh khối trên mặt đất và sự đa dạng loài. Khi so sánh các mô hình khả dụng khác nhau theo bốn tiêu chí AIC, BIC, R2 và MSE, mô hình tốt nhất có dạng như sau:
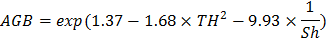 (4)
(4)
Kết quả phân tích độ nhạy của mô hình (4) cho thấy sự đa dạng loài càng cao, sinh khối càng thấp (Hình 4). Vì quá trình tích lũy carbon có nguồn lấy từ sinh khối cây trên mặt đất nên sự đa dạng loài sẽ có tác động tiêu cực tương tự đối với lượng C tích lũy và lượng CO2 hấp thụ của cây keo tai tượng.
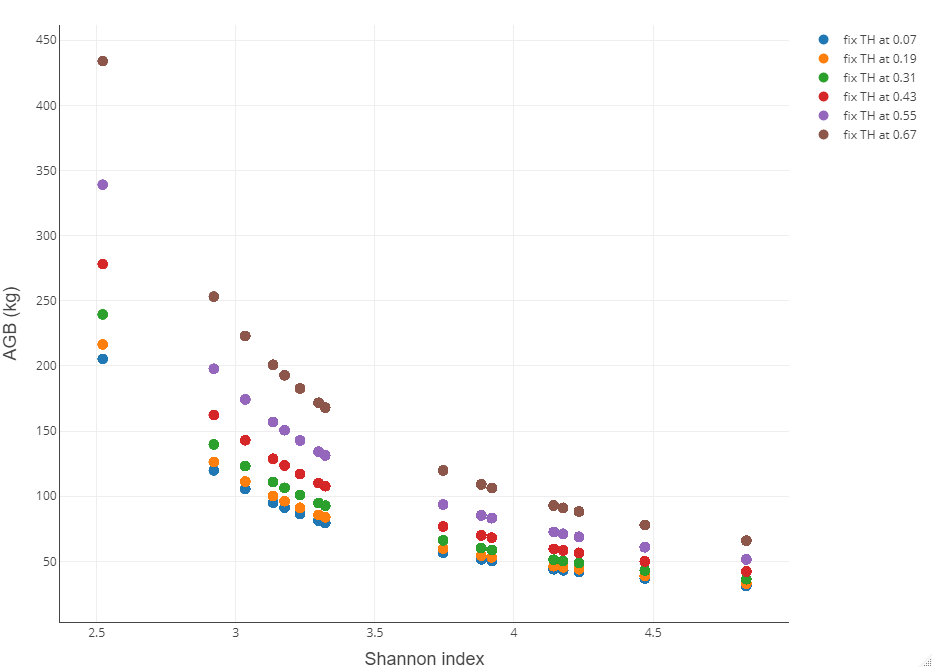
Hình 4. Biểu đồ phân tích độ nhạy của mô hình (4)
Kết quả phân tích độ nhạy mô Hình (4) của nghiên đối với loài keo tai tượng cho thấy rằng sự phong phú và đa dạng của loài có ảnh hưởng tiêu cực đến khả năng tích lũy sinh khối của cây và sự hấp thụ CO2. Đối lập với kết quả này, nghiên cứu của Shuaifeng Li và cộng sự [8] chỉ ra mối quan hệ tích cực giữa sự phong phú về loài và sinh khối trên mặt đất trong rừng thông nguyên sinh. Một nghiên cứu từ Riofrio và cộng sự, trong đó điều tra về tác động của sự kết hợp các loài khác nhau trong một lâm phần cụ thể (Pinus sylvestris L. và Pinus pinaster Ait.) cũng cho thấy rằng việc trồng rừng hỗn giao đã làm thay đổi tích cực sản lượng rừng và sinh khối trên mặt đất [9]. Mối quan hệ tích cực tương tự giữa sinh khối và đa dạng loài cây cũng được tìm thấy khi kiểm tra dữ liệu từ các khu rừng được quản lý, chủ yếu là các lô rừng thứ sinh tương đối non ở Catalonia, Đông Bắc Tây Ban Nha và ở trung bắc Hoa Kỳ [10].
Lưu ý rằng, trong nghiên cứu này, khu vực thu thập số liệu là rừng trồng không được quản lý. Nhiều nhà khoa học đã nghiên cứu rằng, tùy thuộc vào các điều kiện khác nhau của khu rừng, cấu trúc rừng, thành phần loài, tuổi cây,… rừng không được quản lý có thể lưu trữ một lượng carbon lớn hơn một khu rừng đang được quản lý. Tại Việt Nam, mối quan hệ tích cực giữa sự đa dạng loài và sinh khối cây hay nồng độ carbon sẽ có tác động quan trọng đến việc đề xuất và thực hiện chính sách quản lý rừng vì có thể mang lại lợi ích lớn cho bảo tồn đa dạng sinh học còn các trường hợp tương quan tiêu cực thì ngược lại. Tuy nhiên, nâng cao trữ lượng carbon thường không phải là mục tiêu duy nhất trong quản lý rừng. Các nhà quản lý cũng tính đến phát triển các dịch vụ hệ sinh thái và các sản phẩm khác được cung cấp bởi các hệ thống rừng khi xem xét các chế độ, chính sách quản lý khác nhau, vì vậy các lâm phần được quản lý tốt có thể tích lũy nhiều carbon như các lâm phần không được quản lý.
4. Kết luận
Mối quan hệ nghịch giữa sinh khối cây trên mặt đất và sự đa dạng loài thu được từ kết quả phân tích độ nhạy của mô hình tối ưu. Mối quan hệ như vậy không phổ biến khi so sánh với các nghiên cứu khác và sẽ dẫn đến nhiều sự đánh đổi giữa bảo tồn đa dạng sinh học và tích lũy sinh khối hay trữ lượng carbon.
Lời cảm ơn: Các tác giả xin chân thành cảm ơn sự tài trợ kinh phí từ đề tài cấp cơ sở TN.21.18 của Trường Đại học Khoa học Tự nhiên, Đại học Quốc gia Hà Nội và dự án BioEcon (https://bioecon.eu/) để thu thập tài liệu và thực hiện đo đạc lấy số liệu phục vụ cho nghiên cứu.
Đoàn Thị Nhật Minh1,2, Nguyễn Cường Diện1 , Bùi Quang Thành1, Trần Văn Thụy1, Vũ Văn Mạnh1
1 Khoa Môi trường, Trường Đại học Khoa học Tự nhiên, Đại học Quốc gia Hà Nội
2 Trường Đại học Valladolid, Tây Ban Nha
(Nguồn: Bài đăng trên Tạp chí Môi trường, số Chuyên đề Tiếng việt III/2022)
Tài liệu tham khảo
[1]. D. U. Hooper et al., 2005, “Effects of biodiversity on ecosystem functioning: a consensus of current knowledge,” Ecol Monogr, vol. 75, no. 1, pp. 3 - 35, doi: 10.1890/04-0922.
[2]. J. Riofrío, C. Herrero, J. Grijalva, and F. Bravo, 2015, “Aboveground tree additive biomass models in Ecuadorian highland agroforestry systems,” Biomass Bioenergy, vol. 80, pp. 252-259, doi: 10.1016/j.biombioe.2015.05.026.
[3]. F. Kollmann, Tecnologíadelamaderaysusaplicaciones. Tomo primero. Madrid, 1959.
[4]. Phạm Xuân Thành, 2014, “Biomass allometric equation for Acacia auriculiformis forest in Quang Binh province,” Tạp chí Thông tin Khoa học & Công nghệ Quảng Bình, vol. 3, pp. 42-45.
[5]. S. Traoré et al., 2018, “Stand Structure, Allometric Equations, Biomass and Carbon Sequestration Capacity of Acacia mangium Wild. (Mimosaceae) in Cote d’Ivoire,” Open J For, vol. 08, no. 01, pp. 42–60, doi: 10.4236/ojf.2018.81004.
[6]. W. Ounban, L. Puangchit, and S. Diloksumpun, Jan. 2016, “Development of general biomass allometric equations for Tectona grandis Linn.f. and Eucalyptus camaldulensis Dehnh. plantations in Thailand,” Agriculture and Natural Resources, vol. 50, no. 1, pp. 48–53, doi: 10.1016/j.anres.2015.08.001.
[7]. S. Brown, A. J. R. Gillespie, and A. E. Lugo, 1989, “Biomass Estimation Methods for Tropical Forests with Applications to Forest Inventory Data,” Forest Science, vol. 35, no. 4, pp. 881-902.
[8]. S. Li, J. Su, X. Lang, W. Liu, and G. Ou, 2018, “Positive relationship between species richness and aboveground biomass across forest strata in a primary Pinus kesiya forest,” Sci Rep, no. January, pp. 1-9, doi: 10.1038/s41598-018-20165-y.
[9]. J. Riofrío, M. del Río, and F. Bravo, 2016, “Mixing effects on growth efficiency in mixed pine forests,” Forestry: An International Journal of Forest Research, vol. 90, no. 3, pp. 381-392, doi: 10.1093/forestry/cpw056.
[10]. J. P. Caspersen and S. W. Pacala, 2001, “Successional diversity and forest ecosystem function,” pp. 895 - 903.
|
EFFECT OF TREE DIVERSITY ON BIOMASS IN MIXED PLANTATION IN THACH THAT, HANOI Doan Thi Nhat Minh1,2, Nguyen Cuong Dien1 , Bui Quang Thanh1, Tran Van Thuy1, Vu Van Manh1 1Faculty of Environmental Sciences, VNU University of Science 2 University of Valladolid, Spain ABSTRACT: Afforestation has long been recognized as an effective solution to reducing the rate of CO2 increase in the atmosphere. In addition, the information on forest biomass and carbon concentration plays an important role in sustainable management of forest resources. Most of the plantations is monoculture, however, the trend of planting forests with two or more species is being expanded. The results of this study show that the Marteloscope stores 172.80 tons/ha of aboveground tree biomass and 86.40 tons/ha of carbon with high diversity and high evenness. Although the Marteloscope was confirmed as a plantation, the results from the analysis of the spatial distribution descriptive index show that the trees in the study area tend to have a random distribution. Sensitivity analysis indicates that above ground biomass and carbon concentration decrease as the species diversity increase. Key words: Marteloscope, forest structure, sensitivity analysis, forest biomass. |