21/11/2023
Tóm tắt:
Môi trường nuôi cấy ảnh hưởng trực tiếp đến hiệu quả nuôi cấy. Bên cạnh đó, chi phí môi trường còn chiếm đến 50% toàn bộ chi phí sản xuất. Nghiên cứu này thực hiện khảo sát các môi trường nhân tạo thường dùng và thành phần hữu cơ bổ sung từ phân bón thương mại, có hiệu quả tích cực đến sinh khối tảo Chlorella, được phân lập tại Cần Giờ. Trong nghiên cứu này, chúng tôi thực hiện nuôi cấy tảo Chlorella trên 4 loại môi trường (HAMGM, M-8, BG-11 và LC Oligo). Môi trường HAMGM cho kết quả sinh khối khô cao nhất (4,56g/L - ngày 14) và thấp nhất là môi trường LC Oligo (2,29g/L - ngày 14). Nguồn phân bón hữu cơ thương mại như: phân trùn quế (ViF-Maxx), phân bánh dầu (ViF-Mix) và phân bón lá (Terra Sorb 4 Root) được bổ sung vào môi trường HAMGM (1,5g/L) cho hiệu quả tích cực đến khả năng tăng trưởng của tảo Chlorella. Trong đó, phân trùn quế là nguồn dinh dưỡng bổ sung có hiệu quả thúc đẩy tăng trưởng của Chlorella (6,86g/L) cao nhất. Môi trường HAMGM và nguồn dinh dưỡng bổ sung là phân trùn quế, cho thấy có hiệu quả tích cực đến năng suất sinh khối tảo Chlorella.
Nhận bài: 3/8/2023; Sửa chữa: 25/9/2023; Duyệt đăng: 28/9/2023.
Isolation, identification and investigation of the effects of artificial media and commercial organic fertilizers on the growth of microalgae Chlorella in Can Gio
Abstract:
Cultivation efficiency is directly influenced by culture medium, and, medium makes up of 50% total production cost. Our study investigated commonly used artifical medium with organic supplment from commercial organic fertilizer which can be beneficial for Chlorella biomass growth, which was obtained and isolated from Can Gio. In this study, we cultivated Chlorella on 4 medium (HAMGM, M-8, BG-11, and LC Oligo), amongst which the HAMGM medium yielded highest dried biomass (4.56g/L - day 14) and LC Oligo yielded lowest (2.29g/L - day 14). Supplement (at 1.5g/L) from commercial organic fertilizer such as vermicompost (ViF-Maxx), oil cake fertilizer (ViF-Mix), and foilar fertilizer (Terra Sorb 4 Root) all improved Chlorella growth, amongst which vermicompost was the most effective in increasing Chlorella growth (6,86g/L). The HAMGM medium with vermicompost as supplement was shown to be effective in improvement of Chlorella sp. biomass productivity.
Keywords: Chlorella, dried biomass, commercial organic fertilizers.
JEL Classifications: N55, O13, Q15.
1. Đặt vấn đề
Sinh khối vi tảo Chlorella được biết đến với khả năng ứng dụng đa dạng trong đời sống thực tiễn. Tuy nhiên, sản xuất sinh khối tảo Chlorella không được thành công như tảo Spirulina, bởi vì khó khăn trong nuôi cấy và thu hoạch. Trong nuôi cấy sản xuất sinh khối tảo, môi trường nuôi cấy có ảnh hưởng lớn đến hiệu quả sinh khối thu được của toàn bộ quá trình nuôi cấy. Các môi trường khác nhau sẽ có hiệu suất sinh khối thu được khác nhau từ 0,95 - 5 g/L [1]. Chi phí cho môi trường nuôi cấy còn chiếm 50% toàn bộ chi phí sản xuất [2]. Môi trường nuôi cấy phù hợp có vai trò quyết định đến hiệu quả sinh khối vi tảo thu được.
Ngoài ra, một số thành phần hữu cơ như: sodium acetate (nguồn carbon hữu cơ), dịch thuỷ phân chất thải gia súc, và amino acid tự do khi được bổ sung vào trong môi trường nuôi cấy có vai trò nâng cao hiệu quả sinh khối của tảo Chlorella. Môi trường nhân tạo bổ sung thêm nước thải chăn nuôi bò sữa có thể làm tăng hiệu suất sinh khối khô của tảo Chlorella vulgaris lên 0,33 g/L. Amino acid tự do có khả năng thúc đẩy sư tăng trưởng của chủng tảo Chlorella lên đến 110,2% [3]. Vi tảo có thể hấp thu nguồn nito hữu cơ thông qua con đường NtcA nitrogen transcription regulator, nito hữu cơ sẽ được chuyển hoá thành NH4+, cuối cùng chúng sẽ được gắn vào khung cacbon nhờ vào hoạt động của hai enzym là glutamate synthase (GOGAT) và glutamine synthetase (GS) [4]. Tuy nhiên, các nguồn dinh dưỡng hữu cơ như nước thải chăn nuôi hay phân thường tồn tại nhiều chủng vi sinh vật khác nhau. Các chủng vi sinh vật này có nguy cơ gây hại đến sức khoẻ con người, một số chủng thường gặp là: Pseudomonas, Acinetobacter, Klebsiella, Aeromonas spp., Escherichia coli và enterococci [5]. Nguồn phân bón hữu cơ có thành phần tương tự như dịch thuỷ phân phân gia súc, có cả amino acid, và một ưu điểm hơn hết là không có chứa nhóm vi sinh vật nguy hại kể trên.
Việc lựa chọn môi trường nhân tạo phù hợp để nuôi cấy tảo Chlorella là yếu tố quan trọng quyết định năng suất nuôi cấy sinh khối tảo. Ngoài ra, nguồn dinh dưỡng hữu cơ bổ sung đến từ phân bón hữu cơ thương mại có tác động tích cực đến hiệu suất sinh khối tảo, tương tự như các nghiên cứu sử dụng nguồn dinh dưỡng hữu cơ đến từ chất thải chăn nuôi. Nâng cao lượng sinh khối thu được từ 1 lít môi trường nuôi cấy sẽ góp phần làm giảm chi phí sản xuất sinh khối tảo.
2.1. Vật liệu
Mẫu nước có tảo được thu từ thủy vực tạm thời tại Cần Giờ (10.606304, 10.819515 A). Mẫu được giữ trong ống ly tâm 1mL. Mẫu được lọc qua màng lọc có kích thước lỗ 50µm được nuôi trong điều kiện ánh sáng 50 μmol photon.m-2.s-1 và ở nhiệt độ 25oC. Tế bào tảo đơn được hút bằng micropipette và nuôi trong eppendorf 0,2ml chứa 50 µl môi trường BG-11, nuôi cấy ở 50 μmol photon.m-2.s-1 (nguồn sáng đèn huỳnh quang), chu kỳ sáng: 12 sáng/12 tối và ở nhiệt độ 25oC. Các mẫu có hình thái không đồng nhất sẽ được loại bỏ. Mẫu tảo phân lập được ký hiệu là CS.
Môi trường nhân tạo dùng trong thí nghiệm BG-11, LC Oligo, Highly Assimilable Minimal Growth Medium (HAMGM), M-8. Phân bón hữu cơ thương mại dạng lỏng dùng trong thí nghiệm là phân bón lá Terra Sorb 4 Root (amino acid 1,8%), phân trùn quế ViF-Max (đạm tổng 3%), phân bánh dầu thủy phân ViF- Mix (đạm tổng 2%). Lượng phân bón hữu cơ được bổ sung với nồng độ 1,5g.L-1 môi trường HAMGM. Muối Sodium acetate [6] được sử dụng ở nồng độ 1,5g/L môi trường.
2.2. Phương pháp định danh hình thái và phân tử
Tế bào tảo được thu bằng cách ly tâm ở 3.000g trong vòng 5 phút, cặn tảo sẽ được rửa bằng nước cất vô trùng 3 lần. Hình thái tảo sẽ được quan sát dưới kính hiển vi quang học để xác định hình thái. Đối với phương pháp định danh phân tử, DNA tổng số của tảo được tách chiết bằng bộ kit DNeasy Plant Mini (Qiagen, Hilden, Germany). Các vùng trình tự 18S rRNA, rbcL và ITS của chủng tảo phân lập sẽ được đem gửi giải trình tự tại Công ty 1st Base (Malaysia). Cây phát sinh loài sẽ được xây dựng và phân tích thông qua phần mềm MEGA X.
2.3 Phương pháp nuôi cấy vi tảo
Thí nghiệm khảo sát sự ảnh hưởng của môi trường nuôi cấy của Chlorella vulgaris được thực hiện trên hệ thống dịch treo có bổ sung khí (1% CO2 w/w). Các thông số cơ bản cho thí nghiệm bao gồm: Bình nuôi cấy là erlen có thể tích 2 lít (thể tích hoạt động là 1,5 lít), hệ thống bổ sung khí được lọc qua hai hệ thống lọc (màng lọc thô có kích thước 50 µm, màng lọc FINETECH - Đài Loan có kích thước lỗ 0,45 µm). Nút đậy bình erlen có hai ống thủy tinh để dẫn khí. Thí nghiệm sẽ thực hiện trong vòng 14 ngày, chu kỳ chiếu sáng là 12 sáng và 12 tối. Nguồn chiếu sáng trong thí nghiệm được cung cấp bằng đèn LED trắng, mật độ photon quang hợp được duy trì trong khoảng 100 µmol photon.m-2.s-1.
2.4. Phương pháp xác định hàm lượng sinh khối khô
Sinh khối khô tảo thu được bằng cách hút 10ml dịch nuôi cấy qua giấy lọc Whatman (sấy khô ở 100oC đến khối lượng không đổi có giá trị Wf). Sau khi quá trình lọc hoàn tất, tiếp tục sấy khô giấy lọc đến khối lượng không đổi (giá trị Wt). Sinh khối tảo khô được tính theo công thức:
B = Wt – Wf
Trong đó, B: lượng sinh khối khô (g), Wt: tổng khối lượng giấy lọc và sinh khối, Wf: khối lượng giấy lọc.
2.5. Phương pháp xác định hàm lượng Chlorophyll a
10 ml dịch nuôi cấy được ly tâm ở 10.000rpm/phút (5 phút) thu sinh khối và loại bỏ toàn bộ dịch nổi. Phần sinh khối này được bổ sung 2ml acetone 90% và 3 viên bi thủy tinh (d = 5mm) vào ống eppendorf, đem lắc ở vận tốc 300 vòng/phút (15 phút). Quá trình trên được thực hiện cho đến khi dung dịch Acetone không thay đổi màu. Sau cùng dịch tách chiết sẽ được đo ở các bước sóng 750 nm, 664 nm, 647 nm, 630 nm bằng máy đo hấp thu quang phổ (Spectro SC, LaboMed). Hàm lượng chlorophyll a được xác định bằng công thức:
Chlorophyll a (mg/L) = [11,85.(E664 - E750) - 1,54.(E647 - E750) - 0,08.(E630 - E750)].Ve/LVf
E750, E664, E647, E630 : Là giá trị độ hấp thụ lần lượt ở các bước sóng 750 nm ,664 nm, 647 nm, 630 nm, Ve: thể tích Acetone tính bằng lít; Vf: thể tích lọc tính bằng lít; L: đường sáng của cuvet (cm) [8].
Các thí nghiệm được lặp lại 3 lần, số liệu thí nghiệm được xử lý thống kê bằng phần mềm Excel 2019 và SPSS.
3. Kết quả và thảo luận
3.1 Định danh hình thái và định danh phân tử loài tảo phân lập
Kết quả giải trình tự cho thấy, kết quả vùng 18S rRNA (độ bao phủ 100%, độ đồng nhất 100%), ITS (độ bao phủ 100%, độ đồng nhất 100%) và rbcL (độ bao phủ 85,55%, độ đồng nhất 98%) có độ tương đồng cao với cơ sở dữ liệu của Chlorella sorokiniana. Bên cạnh đó, cây phát sinh chủng loài được xây dựng dựa trên 3 marker này được thể hiện qua 2 hình (Hình 1 và Hình 2).

Hình 1. Cây phát sinh chủng loài giản lược mô tả chi tiết vị trí phân bố của chủng vi tảo Chlorella sorokiniana được xây dựng từ sự kết hợp
vùng trình tự 18S - ITS
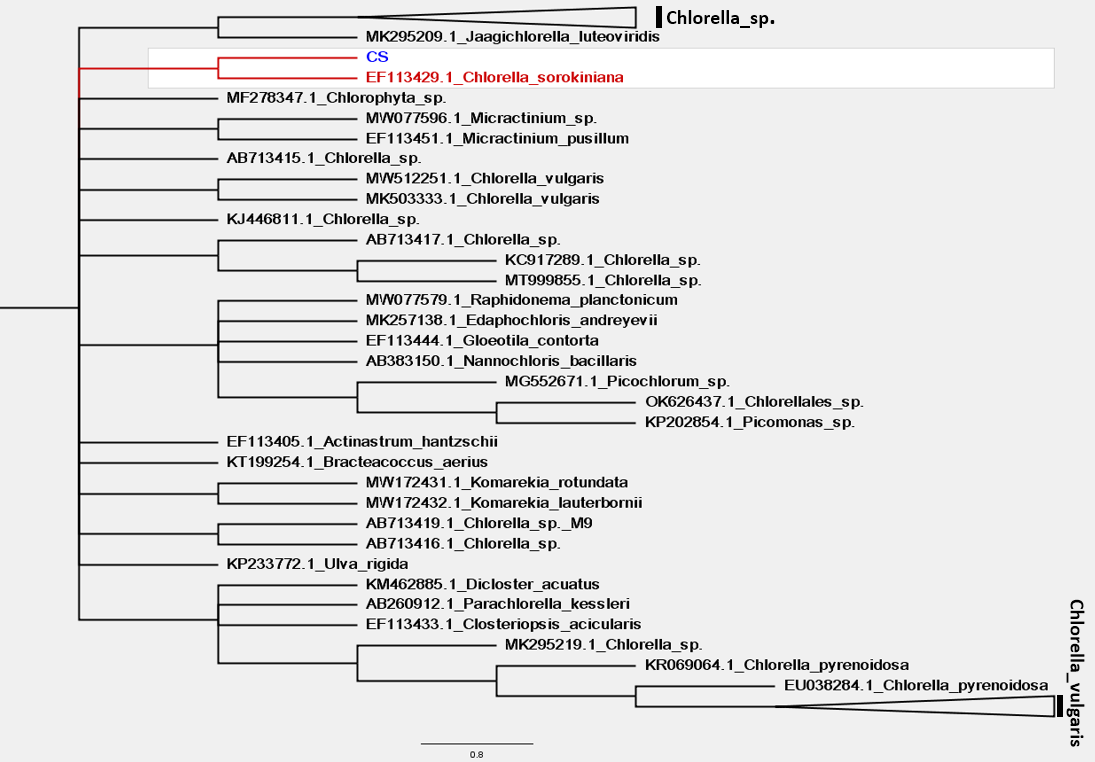
Hình 2. Cây phát sinh chủng loài giản lược mô tả chi tiết vị trí phân bố của chủng vi tảo Chlorella sorokiniana được xây dựng từ vùng trình tự gen rbcL
Kết quả giải trình tự và so sánh trên GenBank của 3 marker phân tử (18S rRNA, ITS và rbcL) dùng trong nghiên cứu này cho thấy, cả 3 vùng đều có sự tương đồng cao với các vùng trình tự đã công bố của chủng Chlorella sorokiniana.
Cây phát sinh chủng loài ở trình tự hai vùng 18S và ITS cho thấy, tảo phân lập được trong nghiên cứu này nằm cùng nhóm với chủng tảo Chlorella sorokiniana (giá trị bootstrap lớn hơn 50%). Bên cạnh đó, kết quả xây dựng cây phát sinh chủng loài dựa trên vùng trình tự rbcL cũng cho kết quả tương tự là chủng tảo phân lập được trong nghiên cứu này đều nằm chung nhóm với chủng Chlorella sorokiniana. Kết quả hình thái và cây phát sinh chủng loài cho thấy, chủng tảo phân lập được có độ tương đồng cao với chủng Chlorella sorokiniana.
3.2. Sự tăng trưởng của tảo Chlorella sorokiniana trong các môi trường nhân tạo
Nghiên cứu thực hiện khảo sát khả năng tăng trưởng của tảo Chlorella sorokiniana ở các môi trường nhân tạo trong vòng 14 ngày. Kết quả Hình 3 cho thấy, cả 4 môi trường nuôi cấy đều cho kết quả sinh khối khô ở ngày thứ 14 đều cao hơn 2g/L. Hàm lượng Chlorophyll a của tảo ở ngày thứ 14 cho thấy, chưa có dấu hiệu giảm. Hàm lượng sinh khối khô sau 14 ngày nuôi cấy cao nhất ở môi trường HAMGM (4,56g/L). Đối với môi trường BG-11, kết quả trong nghiên cứu này (2,64g/L) cho thấy cao hơn các nghiên cứu trước đây đã công bố của Chan Yoo (1,46g/L) [7], Ghulam Mujtaba (1,9g/L) [10]. Tuy nhiên, hàm lượng sinh khối khô của nghiên cứu này thấp hơn so với công bố của Sigita (2,8g/L) [11]. Nguyên nhân dẫn đến việc thấp hơn này do phương pháp nuôi cấy. Tảo ở nghiên cứu của Sigita được thực hiện trên hệ thống quang sinh học hình trụ (cylindrical glass) và nuôi cấy bán liên tục. Môi trường mới được thay và bổ sung lên đến 50% thể tích nuôi cấy. Ngoài ra, các yếu tố như mật độ photon chiếu sáng và nhiệt độ nuôi cấy cũng có khác biệt (150 µmol photon.m2s-1, 30 oC, 2% CO2). Vi tảo trong môi trường thiếu hụt nitrogen, quá trình sinh dưỡng của tảo sẽ chuyển sang dạng tích lũy lipid và hàm lượng Chlorophyll a có xu hướng giảm dần. Ngoài ra, khi môi trường thiếu hụt nitrogen, lipid được tổng hợp chủ yếu ở dạng neutral lipid [11]. Chlorella pyrenoidosa được nuôi cấy trong môi trường dinh dưỡng đầy đủ sẽ có hàm lượng polyunsaturated fatty acid và unsaturated fatty acid cao hơn khi ở điều kiện hoàn cảnh bất lợi [12]. Với mục đích ứng dụng sinh khối tảo để làm nguồn thức ăn có giá trị dinh dưỡng cao. Do đó, nghiên cứu này không tiếp tục thực hiện theo dõi khảo sát sau 14 ngày.
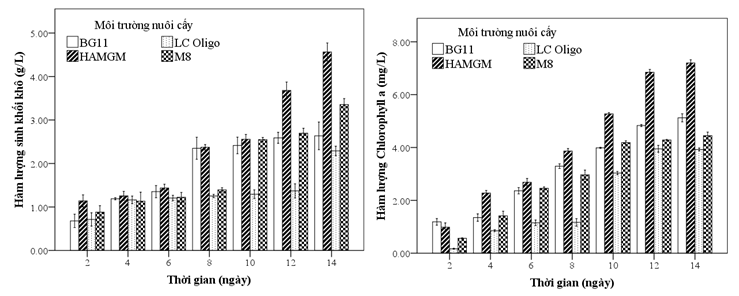
Hình 3: Hàm lượng sinh khối khô và chlorophyll a của Chlorella sorokiniana
BG11: Chlorella sorokiniana nuôi cấy ở môi trường BG-11.
LC Oligo: Chlorella sorokiniana nuôi cấy ở môi trường LC Oligo.
HAMGM: Chlorella sorokiniana nuôi cấy ở môi trường HAMGM.
M-8: Chlorella sorokiniana nuôi cấy ở ở môi trường M-8.
3.3. Sự tăng trưởng của tảo Chlorella sorokiniana ở môi trường nhân tạo bổ sung các nguồn phân bón hữu cơ
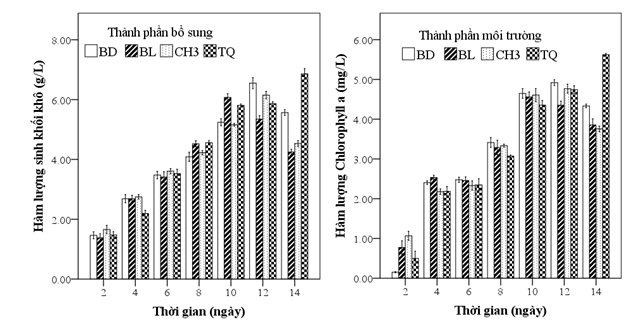
Hình 4: Hàm lượng sinh khối khô và Chlorophyll a của Chlorella sorokiniana
BD: môi trường bổ sung phân bánh dầu thủy phân ViF- Mix
BL: Môi trường bổ sung phân bón lá Terra Sorb 4 Root
CH3: Môi trường bổ sung muối CH3COONa
TQ: Môi trường bổ sung phân trùn quế ViF-Max
Ở thí nghiệm này, vi tảo được nuôi cấy ở môi trường HAMGM và bổ sung thêm các nguồn phân bón hữu cơ. Kết quả Hình 4 cho thấy, sinh khối khô tảo ở các nghiệm thức cao nhất ở môi trường bổ sung phân trùn quế (6,86g/L, ngày 14). Hàm lượng Chlorophyll a của tảo tăng đều trong giai đoạn từ 2 - 10 ngày; từ ngày thứ 12, hàm lượng ở môi trường bổ sung phân bón lá giảm, trong khi đó ở 3 môi trường còn lại tăng. Ngày thứ 14, môi trường trùn quế đạt hàm lượng cao nhất, 3 môi trường còn lại thì giảm.
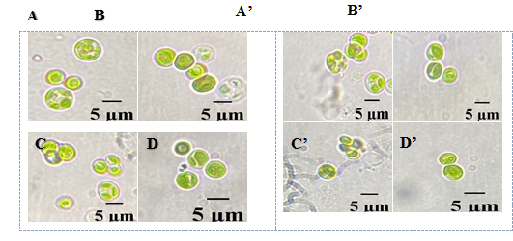
Hình 5. Tảo Chlorella sorokiniana nuôi cấy ở môi trường HAMGM và bổ sung thêm các nguồn phân bón hữu cơ
A (ngày đầu) - A’ (sau 14 ngày): Hình thái tảo Chlorella sorokiniana ở môi trường bổ sung ViF- Mix.
B (ngày đầu) - B’ (sau 14 ngày): Hình thái tảo Chlorella sorokiniana ở môi trường bổ sung Terra Sorb 4 Root.
C (ngày đầu) - C’ (sau 14 ngày): Hình thái tảo Chlorella sorokiniana ở môi trường bổ sung CH3COONa.
D (ngày đầu) - D’ (sau 14 ngày): Hình thái tảo Chlorella sorokiniana ở môi trường bổ sung ViF-Max.
Kết quả sinh khối cao nhất đạt được ở môi trường bổ sung phân trùn quế cao gấp 2,4 lần khi so với các nghiên cứu nuôi cấy trong môi trường nước thải nuôi trồng thủy sản (2,858g/L) [13]. Tảo Chlorella nuôi cấy trong hệ thống quang sinh học Airlift với nguồn phân bón thương mại (nguồn nitrogen từ urea và ammonia) cũng cho kết quả thấp hơn (ngày 14 là 1,988g/L) [14].
Các nghiên cứu về sự ảnh hưởng của các loại amino acid có sự ảnh hưởng đến khả năng tăng trưởng của vi tảo Chlorella. Một nghiên cứu gần đây cho thấy, aspartic acid và arginine được cho rằng có ảnh hưởng tích cực đến khả năng tăng trưởng của Chlorella; Bổ sung aspartic acid có khả năng tăng cường hiệu suất sinh khối khô lên đến 110%, còn arginine thì chỉ ở mức 62%; Glucose, tryptophan, alanine được cho là các thành phần có ảnh hưởng đến khả năng sinh trưởng, cũng như tổng hợp lipid của tảo Chlorella sorokiniana [15]. Trong sản xuất các loại phân bón lá, một số chất kích thích sinh trưởng thường được phép bổ sung nhằm thúc đẩy sinh trưởng hoặc thúc đẩy ra hoa (Thông tư số 36/2010/TT-BNNPTNT của Bộ Nông nghiệp và Phát triển nông thôn về việc ban hành quy định sản xuất, kinh doanh và sử dụng phân bón). Các chất kích thích sinh trưởng thuộc nhóm auxin làm tăng hàm lượng Chlorophyll a và b lên đến 213 - 273%, carotenoid tăng 164 - 258% [15]. Các chất kích thích sinh trưởng thuộc nhóm cytokinin có thể thúc đẩy tốc độ phân chia của tảo Chlorella pyrenoidosa lên 12-16%, đồng thời tăng cường hàm lượng chlorophyll của tảo lên 183-256% [17]. Gibberellins thì có khả năng tăng cường sinh khối khô Chlorella [17]. Abscisic Acid (ABA) giúp tăng cường hấp thu nitrogen vào trong tế bào tảo [18]. Hỗn hợp NAA, gibberellins GA3 và zeatin có thể kích thích làm tăng hàm lượng sinh khối khô lên 170% [19]. Đây có thể là một phần nguyên nhân gây ra sự khác biệt giữa các loại phân bón khảo sát bên cạnh sự ảnh hưởng của nguồn nitrogen hữu cơ.
4. Kết luận
Trong 4 môi trường nhân tạo khảo sát, môi trường nhân tạo HAMGM cho kết quả sinh khối khô tốt nhất. Phân bón hữu cơ bổ sung thêm vào môi trường nhân tạo có khả năng thúc đẩy tăng trưởng của tảo C. sorokiniana, trong đó dịch thủy phân trùn quế cho kết quả tăng trưởng tốt nhất trong các loại phân bón khảo sát.
Lời cảm ơn: Nghiên cứu này tài trợ bởi Đại học Nguyễn Tất Thành trong khuôn khổ nhiệm vụ Đề tài cấp trường 2020.01.154.
Phan Minh Tâm1*, Bùi Thị Mỹ Ngọc, Lê Tuấn Lộc4,
Ông Bỉnh Nguyên4, Đỗ Thị Thu Huyền3, Huỳnh Văn Biết1,2
1 Khoa Khoa học Sinh học, Trường Đại học Nông Lâm TP. Hồ Chí Minh (HCM)
2 Viện nghiên cứu Công nghệ Sinh học và Môi trường
Trường Đại học Nông Lâm TP. HCM
3Viện Môi Trường và Tài Nguyên, Đại học Quốc Gia TP. HCM
4 Viện Kỹ thuật Công nghệ cao NTT, Trường Đại học Nguyễn Tất Thành
(Nguồn: Bài đăng trên Tạp chí Môi trường, số Chuyên đề Tiếng Việt III/2023)
Tài liệu tham khảo
[1]. Weiguo Zhang, Effects of various amino acids as organic nitrogen sources on the growth and biochemical composition of Chlorella pyrenoidosa, Bioresource technology, 197: 458 - 464, 2015.
[2]. Ao Xia, Innovation in biological production and upgrading of methane and hydrogen for use as gaseous transport biofuel, Biotechnology advances, 34(5): 451-472, 2016.
[3]. Yeh, K. L., & Chang, J. S. Effects of cultivation conditions and media composition on cell growth and lipid productivity of indigenous microalga Chlorella vulgaris ESP-31. Bioresource technology, 105, 120 - 127, 2012.
[4]. Dagon Manoel Ribeiro,, Effect of different culture media on growth of Chlorella sorokiniana and the influence of microalgal effluents on the germination of lettuce seeds, J Appl Biol Biotechnol, 7(1): 6 - 10, 2019.
[5]. Amaro, H. M., Microalgae systems-environmental agents for wastewater treatment and further potential biomass valorisation. Journal of Environmental Management, 337, 117678, 2023.
[6]. Pang, N., Effects of C5 organic carbon and light on growth and cell activity of Haematococcus pluvialis under mixotrophic conditions. Algal research, 21, 227 - 235, 2017.
[7]. Merve Tunali, Effect of microplastics and microplastic-metal combinations on growth and chlorophyll a concentration of Chlorella vulgaris, Science of the Total Environment, 743: 140479, 2020.
[8]. Ritchie, R. J, Universal chlorophyll equations for estimating chlorophylls a, b, c, and d and total chlorophylls in natural assemblages of photosynthetic organisms using acetone, methanol, or ethanol solvents, Photosynthetica, 46, 115 - 126, 2008.
[9]. Ghulam Mujtaba, Lipid production by Chlorella vulgaris after a shift from nutrient-rich to nitrogen starvation conditions, Bioresource technology, 123: 279 - 283, 2012.
[10]. Sigita Vaičiulytė, Batch growth of Chlorella vulgaris CCALA 896 versus semi-continuous regimen for enhancing oil-rich biomass productivity, Energies, 7(6): 3840 - 3857, 2014.
[11]. Ma, X. N., Lipid production from Nannochloropsis. Marine drugs, 14(4), 61, 2016.
[12]. Dong, L., Li, D., Characteristics of lipid biosynthesis of Chlorella pyrenoidosa under stress conditions. Bioprocess and biosystems engineering, 43(5), 877 - 884, 2020.
[13]. Kulwa Mtaki, Supplementing wastewater with NPK fertilizer as a cheap source of nutrients in cultivating live food (Chlorella vulgaris), Annals of Microbiology, 71(1): 1 - 13, 2021.
[14]. Saad Hanash Ammar, Cultivation of Microalgae Chlorella vulgaris in Airlift photobioreactor for Biomass Production using commercial NPK Nutrients, Al-Khwarizmi Engineering Journal, 12(1): 90-99, 2016.
[15]. Ngang kham, M., Biochemical modulation of growth, lipid quality and productivity in mixotrophic cultures of Chlorella sorokiniana. Springer Plus, 1(1), 1 - 13, 2012.
[16]. Czerpak R, Effect of auxin precursors and chemical analogues on the growth and chemical composition in Chlorella pyrenoidosa Chick. Acta Soc Bot Pol 63:279 - 286, 1994.
[17]. Piotrowska A, Cellular response of light/dark- grown green alga Chlorella vulgaris Beijerinck (Chlorophyceae) to exogenous adenine- and phenylurea-type cytokinins. Acta Physiol Plant 31:573 - 585, 2009.
[18]. Ulrich WR, Effect of abscisic acid on nitrogen uptake, respiration and photosynthesis in green algae. Plant Sci Lett 37:9 - 14, 1984.
[19]. Hunt RW, Effect of biochemical stimulants on biomass productivity and metabolite content of the microalga, Chlorella sorokiniana. Appl Biochem Biotechnol 162:2400 - 2414, 2010.