

06/05/2024
Tóm tắt:
Việc ứng dụng các chủng vi sinh vật (VSV) hữu ích cũng như các sản phẩm thứ cấp của chúng trong chuỗi sản xuất chăn nuôi, trồng trọt, công nghiệp thực phẩm tuần hoàn, xanh-bền vững đang được phát triển rộng rãi. Nghiên cứu này nhằm mục đích tuyển chọn được các chủng nấm nội sinh trên cây Thạch tùng javanica Việt Nam có khả năng sinh đa enzym ngoại bào và đa kháng các VSV gây bệnh bằng phương pháp xác định khả năng sinh enzym ngoại bào của các chủng vi nấm và khuếch tán trên đĩa thạch. Kết quả cho thấy, 9 chủng nghiên cứu đều có khả năng sinh từ 1 - 5 loại enzym và ức chế từ 1 - 5 loài VSV gây bệnh với hoạt tính tiềm năng; 2 chủng TLC11 và TLC9 sinh 4 ÷ 5 enzym (cellulase, lipase, protease, phosphatase, β-galactosidase) với đường kính vòng thủy phân cao nhất 22 ÷ 25 mm (protease) và 20 ÷ 23 mm (lipase); chủng TLC13 ức chế cả 5 chủng VSV kiểm định (Escherichia coli, Staphylococcus aureus, Candida albican, Bacillus cereus, Pseudomonas aerigunosa) với hoạt tính cao nhất với 3 loài B. cereus (24 ± 1,2 mm), P. aerigunosa (26 ± 1,1 mm) và C. albican (36 ± 1,5 mm); 2 chủng TLC10 và TLC19 kháng 4/5 VSV kiểm định ngoại trừ S. aureus (TLC10) và E. coli (TLC19). Các chủng này sẽ là nguồn nguyên liệu tiềm năng trong các nghiên cứu sâu hơn nhằm hướng tới có thể ứng dụng xa hơn trong các lĩnh vực sản xuất nông-công-ngư nghiệp bền vững.
Từ khóa: Hoạt tính sinh học, Thạch tùng javanica, vi nấm nội sinh, enzym ngoại bào, đối kháng vi sinh vật.
Ngày nhận bài: 14/3/2024 ; Ngày sửa chữa: 2/4/2024; Ngày duyệt đăng: 12/4/2024
Biologycal activity research of endophytic fungi on Huperzia javanica plant with application orientation in the circular economy
The application of beneficial microorganisms as well as their secondary products in the livestock, agriculture, and food industry supply chains for sustainable green production is a developing trend. The research direction of selecting multi-activity strains to enhance the effectiveness of practical applications has been receiving attention. This study aims to select endophytic fungal strains on Huperzia javanica plant in Vietnam that have the ability to produce multi-extracellular enzymes and multi-resistance to pathogenic microorganisms by determining enzyme activity and testing antimicrobial activity. The results showed the following: (1) All 9 strains have the ability to produce 1 to 5 types of enzymes and inhibit 1 to 5 pathogenic microorganisms with potential activities; Strains TLC11 and TLC9 produce 4÷5 enzymes (cellulase, lipase, protease, phosphatase and β-galactosidase) with the highest hydrolysis zone diameters of 22÷25 mm (protease) and 20÷23 mm (lipase); (2) Strain TLC13 inhibits all 5 tested microbial strains Escherichia coli, Staphylococcus aureus, Candida albican, Bacillus cereus, Pseudomonas aerigunosa) with the highest activity against 3 species B. cereus (24±1.2 mm), P. aerigunosa ( 26±1.1 mm) and C. albican (36±1.5 mm); Strains TLC10 and TLC19 are resistant against 4/5 tested microorganisms except S. aureus (TLC10) and E. coli (TLC19). These strains could be a potential sources for further in-depth research aiming to expand their applications in sustainable agriculture, aquaculture, and industry production fields.
Keyword: Biological activity, Huperzia javanica, endophytic fungi, extracellular enzyme, antimicrobial.
JEL Classifications: N50, N53, N57, O13.
1. ĐẶT VẤN ĐỀ
Hiện nay, nền kinh tế tuần hoàn cũng như nền kinh tế xanh, tăng trưởng xanh là mô hình kinh tế hướng tới sử dụng hiệu quả, tiết kiệm tài nguyên, tái chế chất thải góp phần mang lại hiệu quả kinh tế và bền vững môi trường. Kinh tế tuần hoàn được xem là xu thế tất yếu của thời đại và là cuộc cách mạng công nghiệp xanh của thế kỷ 21. Theo đó, phát triển nền nông nghiệp - công nghiệp chế biến xanh, bền vững đang được Chính phủ chú trọng phát triển (Quyết định số 687/QĐ-TTg ngày 7/6/2022 của Thủ tướng Chính phủ phê duyệt Đề án Phát triển kinh tế tuần hoàn ở Việt Nam và Quyết định số 882/QĐ-TTg ngày 22/7/2022 của Thủ tướng Chính phủ phê duyệt Kế hoạch hành động quốc gia về tăng trưởng xanh giai đoạn 2021-2030). Việc ứng dụng các chủng VSV hữu ích (vi khuẩn, vi nấm, xạ khuẩn, nấm men…) cũng như các sản phẩm thứ cấp của chúng trong chuỗi sản xuất chăn nuôi, trồng trọt và công nghiệp chế biến thực phẩm tuần hoàn, xanh, bền vững là hướng ứng dụng đang được phát triển rộng rãi (tăng cao sức khỏe, năng suất, chất lượng cây trồng, vật nuôi, sản phẩm sau thu hoạch; xử lý nước-chất thải, phế phụ phẩm nông - công nghiệp thành các sản phẩm hữu ích như phân bón, thức ăn chăn nuôi, vật liệu tự nhiên, nước tưới tiêu…). Trong đó, các chủng VSV có khả năng sinh tổng hợp các enzym ngoại bào (xellulose, proteinase, lipase, amylase…) và kháng vi khuẩn gây bệnh (Escherichia coli, Staphylococcus aureus, Bacillus cereus, Pseudomonas aerigunosa…) như vi khuẩn Bacillus subtillis, B. megaterium, Lactobacillus casei, L. plantarums, Rhodopseudomonas, Azotobacter, Azospirillum, Enterobacter…; xạ khuẩn Streptomyces, Actinomyces...; vi nấm Trichoderma harzianum, Aspergilus tubingensis… đã được sử dụng phổ biến nhằm thay thế, giảm thiểu lượng thức ăn, phân bón, thuốc kháng sinh và các chất hóa học khác (Sindhu và cs, 2018; Inamuddin và cs, 2022).
Tuy nhiên, để tăng cường hiệu quả ứng dụng trong thực tế thì việc nghiên cứu tuyển chọn các chủng đa hoạt tính là cần thiết. Vi nấm nội sinh (VNNS) thực vật (đặc biệt là ở cây thảo dược) đại diện cho một trong những lựa chọn thay thế đầy tiềm năng vì chúng đã chứng tỏ được hiệu quả cao trong việc sản xuất các chất chuyển hóa có hoạt tính sinh học mới, không chỉ có đặc tính kháng khuẩn mà còn có hoạt tính sinh học khác trên phạm vi rộng. Các loài này sống trong các mô và cơ quan khác nhau của cây khỏe mạnh ở một giai đoạn nhất định hoặc tất cả các giai đoạn trong vòng đời của chúng, ngoài khả năng có thể sinh tổng hợp các hợp chất có hoạt tính sinh học tương ứng cây chủ, chúng còn có thể sinh các chất có hoạt tính khác (enzyme, chất kháng khuẩn, potein, alcaloit, polyketit…) có tác dụng giúp cây chủ tăng sinh trưởng, ức chế bệnh tật, chống chịu mặn - hạn hán - nhiệt độ cũng như có thể ứng dụng trong các ngành công nghiệp thực phẩm, nông - ngư nghiệp, môi trường và dược phẩm (Daniel và cs, 2022; Fatima và cs, 2022; Cripwell và cs, 2021; Lu và cs, 2021; Jouda và cs, 2014).
Thạch tùng javanica (H. javanica) là loài cây dược liệu quý (hiện đang trong diện bảo tồn) thuộc họ Thông đất (Lycopodiaceae) được biết đến trong hỗ trợ điều trị một số bệnh về thần kinh, phong tê thấp, viêm gan, tiêu chảy… H. javanica ưa ẩm và bóng mát, mọc trên đất ẩm có tầng dày và nhiều mùn, ở độ cao 1.000-1.500m; phân bố ở Trung Quốc, Ấn Độ, Nhật Bản và Việt Nam (Sun và cs, 2015). Đã có nhiều công bố nghiên cứu về hoạt tính sinh học của các chủng VNNS ở các cây dược liệu khác, tuy nhiên, các nghiên cứu trên cây H. javanica hầu như chưa có. Nghiên cứu này tập trung vào việc khảo sát, tuyển chọn các chủng VNNS cây Thạch tùng javanica Việt Nam có khả năng sinh đa enzyme và đa kháng VSV gây bệnh với mục tiêu hướng tới các nghiên cứu, khai thác ứng dụng xa hơn trong thực tế sản xuất nông - công - ngư nghiệp - dược phẩm tuần hoàn và bền vững, góp phần giảm thiểu ô nhiễm môi trường.
2. VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU
2.1. Vật liệu nghiên cứu
Trong nghiên cứu trước đây, 9 chủng VNNS đã được phân lập và tuyển chọn từ cây Thạch tùng javanica phân bố tại Hà Giang (Việt Nam) có khả năng sinh tổng hợp hoạt chất dược phẩm huperzine (là một alcaloit hỗ trợ trong điều trị bệnh sa sút trí tuệ, đặc biệt là bệnh Alzheimer). Trong nghiên cứu này, các chủng được tiếp tục nghiên cứu sâu hơn về các đặc tính sinh học khác nhằm khảo sát và tuyển chọn được chủng đa sinh tổng hợp enzyme và đa kháng VSV gây bệnh hướng tới các ứng dụng khác trong sản xuất nông - công - ngư nghiệp - dược phẩm tuần hoàn và bền vững. Các chủng bao gồm: Neurospora calospora TLC9, N. calospora TLC10, N. calospora TLC11, Schizophyllum commune TLC12, Epicoccum sorghinum TLC13, Alternaria tenuissima TLC14, Daldinia sp. TLC19, Cephalotrichum sp. TLC20, Schizophyllum sp. TLC22. Các chủng VSV kiểm định: E. coli (ATCC 25922), S. aureus (ATCC 33591), C. albican (ATCC 10231), B. cereus (ATCC 11778), P. aerigunosa (ATCC 27853) do Trung tâm Giống và Bảo tồn nguồn gen vi sinh vật, Viện Công nghệ sinh học cung cấp.
2.2. Phương pháp nghiên cứu
2.2.1. Phương pháp xác định khả năng sinh enzym ngoại bào của các chủng vi nấm
Xác định khả năng sinh amylase
Các chủng vi nấm được nuôi trên môi trường dịch thể PDB ở 28oC trong 5 - 7 ngày. Chuẩn bị đĩa môi trường cơ chất chứa agar 20 g/l có bổ sung 1% tinh bột, khoan các giếng có đường kính 8 mm, mỗi giếng bổ sung 100 µl dịch ngoại bào vi nấm. Đối chứng âm là môi trường PDB không nuôi cấy vi nấm. Giữ đĩa ở 4oC qua đêm cho enzym khuếch tán vào môi trường. Ủ tiếp ở 37ºC trong khoảng 24 giờ để enzym hoạt động. Hoạt độ tương đối của enzym được xác định dựa trên hiệu số D-d (mm). Trong đó D là đường kính vòng phân giải (mm), d là đường kính lỗ thạch (mm). D-d > 25 mm: hoạt tính enzym rất mạnh; D-d = 20-25 mm: hoạt tính enzym mạnh; D-d = 10 - 20 mm: hoạt tính enzym trung bình; D-d < 10 mm: hoạt tính enzym yếu. Thí nghiệm lặp lại 3 lần.
Xác định khả năng sinh protease, xellulase, lipase và phosphatase
Tiến hành tương tự phương pháp xác định khả năng sinh amylase của các chủng vi nấm với môi trường cơ chất bổ sung 1% casein, carboxyl methyl cellulose (CMC), Tributylin và Ca3(PO4)2 để xác định khả năng sinh các enzym tương ứng protease, xellulase, lipase và phosphatase.
Xác định hoạt tính enzym β-galactosidase
X-gal hòa trong dimethyl sulphoxide đạt nồng độ 20µg/mL, bảo quản trong tối ở -20oC. Môi trường PDA sau khi hấp vô trùng được đổ đĩa, sau đó gạt đều 50µl chỉ thị X-gal trên bề mặt môi trường PDA thạch đĩa. Cấy chấm điểm các chủng VNNS trên môi trường thạch đĩa có chỉ thị X-gal. Nuôi ở tủ ấm 28oC trong 3 - 10 ngày, các chủng nấm nội sinh tạo màu xanh da trời trên đĩa chỉ thị là các chủng có khả năng sinh tổng hợp enzym β-galactosidase. Thí nghiệm lặp lại 3 lần.
2.2.2. Phương pháp khuếch tán trên đĩa thạch
Hoạt tính kháng VSV kiểm định được xác định theo phương pháp khuếch tán trên đĩa thạch. Các chủng vi nấm nghiên cứu được nuôi lắc trong môi trường PDB 5-7 ngày, thu dịch chiết nấm. Nhỏ 100µl dịch chiết nấm vào mỗi giếng được tạo trên đĩa môi trường LBA đã được cấy chang VSV kiểm định. Giếng nhỏ môi trường PDB được sử dụng làm đối chứng âm, đối chứng dương là giếng nhỏ kháng sinh ampicllin nồng độ 1 mg/ml. Đĩa được giữ ở 4oC trong 2-4 giờ để enzym khuếch tán vào môi trường, sau đó ủ đĩa ở 37oC trong 24 giờ. Hoạt tính kháng khuẩn được xác định bằng đường kính của vòng vô khuẩn D-d (mm), trong đó: D là đường kính vòng vô khuẩn, d là đường kính giếng. Thí nghiệm lặp lại 3 lần.
3. KẾT QUẢ NGHIÊN CỨU VÀ THẢO LUẬN
3.1. Khả năng sinh enzym của các chủng VNNS
9 chủng VNNS được thử nghiệm khả năng sinh 6 enzym ngoại bào, bao gồm cellulose, lipase, protease, amylase, phosphatase và β-galactosidase.
Khả năng sinh enzym ngoại bào của 9 chủng vi nấm nghiên cứu được thể hiện trong Bảng 1, Hình 1 và Hình 2 cho thấy, tất cả 9 chủng đều có khả năng sinh từ 1 đến 5 loại enzym thử nghiệm; trong đó, số chủng có khả năng sinh lipase và protease chiếm tỷ lệ cao nhất với 7 chủng (77,77%), tiếp đến là số chủng sinh cellulose (6 chủng; 66,66%), số chủng sinh phosphatase chiếm tỷ lệ 33,33% (3 chủng), 2 chủng sinh β-galactosidase (tỷ lệ 22,22%) và chỉ duy nhất 1 chủng TLC10 sinh amylase (11,11%) với hoạt tính thấp. Trong số các enzym do các chủng nghiên cứu sinh ra, protease là hoạt tính được thể hiện mạnh nhất ở các chủng nấm, 3 chủng (TLC9, TLC11 và TLC12) có đường kính vòng thủy phân từ 20 ÷ 25 mm, 4 chủng (TLC13, TLC14, TLC20 và TLC22) đường kính vòng thủy phân trên 25 mm. Trong 7 chủng sinh lipase, 6 chủng (TLC9, TLC10, TLC11, TLC14, TLC19 và TLC22) có hoạt tính lipase mạnh (đường kính vòng thủy phân từ 20 ÷ 25 mm).
Khi nuôi vi nấm trên môi trường có chất chỉ thị X-gal, các chủng bắt màu xanh cho thấy có khả năng sinh β-galactosidase. Chủng có thời gian bắt màu nhanh, có màu đậm sẽ có khả năng sinh β-galactosidase cao hơn. Trong 3 chủng có hoạt tính sinh β-galactosidase, chủng TLC14 có khuẩn lạc chuyển màu xanh trong thời gian hình thành khuẩn lạc rất ngắn (sau 2 ngày nuôi cấy), khả năng bắt màu mạnh chứng tỏ có khả năng sinh β-galactosidase cao. Trong khi đó 2 chủng còn lại (TLC11 và TLC20) khuẩn lạc chuyển màu xanh chậm hơn (sau 3 ngày nuôi cấy) và độ bắt màu của các khuẩn lạc không đồng đều nên có thể có khả năng sinh β-galactosidase không cao.
Bảng 1. Khả năng sinh enzym của các chủng vi nấm nghiên cứu
|
TT |
Chủng |
Hoạt tính enzym (mm) |
|||||
|
Cellulase |
Lipase |
Protease |
Phosphatase |
Amylase |
β-galactosidase |
||
|
1 |
TLC9 |
12 ± 0,9 |
20 ± 1,4 |
25 ± 1,5 |
8 ± 0,7 |
- |
- |
|
2 |
TLC10 |
4 ± 0,6 |
23 ± 1,2 |
- |
- |
5 ± 0,6 |
- |
|
3 |
TLC11 |
9 ± 0,8 |
23 ± 1,1 |
22 ± 1,2 |
6 ± 0,5 |
- |
+ |
|
4 |
TLC12 |
7 ± 0,5 |
- |
22,5 ± 1,2 |
- |
- |
- |
|
5 |
TLC13 |
- |
- |
31 ± 1,6 |
- |
- |
- |
|
6 |
TLC14 |
- |
28 ± 1,7 |
32 ± 1,5 |
- |
- |
+ |
|
7 |
TLC19 |
3 ± 0,3 |
25 ± 1,5 |
- |
3 ± 0,3 |
- |
- |
|
8 |
TLC20 |
- |
10 ± 0,9 |
34 ± 1,7 |
- |
- |
+ |
|
9 |
TLC22 |
11 ± 0,9 |
24,5 ± 1,2 |
28 ± 1,3 |
- |
- |
- |
|
10 |
ĐC (-) |
- |
- |
- |
- |
- |
|
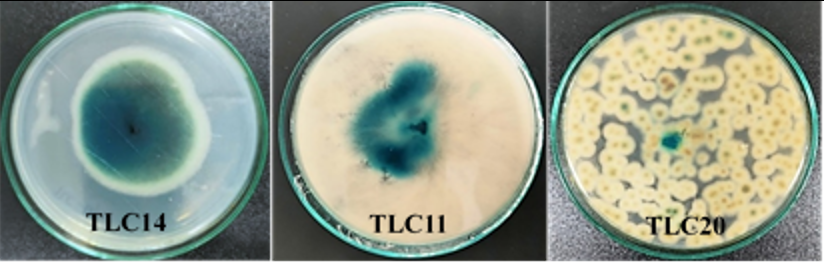
Hình 1. Các chủng nấm có khả năng sinh
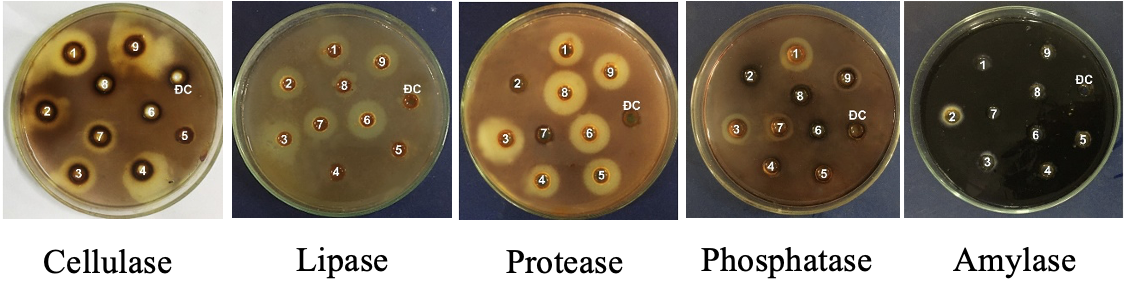
Hình 2. Khả năng sinh tổng hợp enzym của các chủng vi nấm (1. TLC9, 2. TLC10, 3. TLC11, 4. TLC12, 5. TLC13, 6. TLC14, 7. TLC19, 8. TLC20, 9. TLC22, ĐC (-): đối chứng âm)
Trong 6 enzym thử nghiệm, chủng TLC11 cho khả năng sinh 5/6 loại ngoại trừ enzym amylase; tiếp theo là chủng TLC9 sinh 4/5 loại enzym; 4 chủng sinh 3 loại enzym là TLC10, TLC19, TLC20 và TLC22 và chủng TLC13 duy nhất chỉ sinh mạnh 1 loại enzym protease với vòng hoạt lực 31 ± 1,6 mm. Đáng chú ý, 3 chủng TLC9, TLC10 và TLC11 cùng là loài N. calospora nhưng cho khả năng sinh enzym là khác nhau.
Theo các nghiên cứu trước đây, enzym là chất xúc tác sinh học hơn 5.000 loại phản ứng hóa sinh giúp thúc đẩy quá trình trao đổi chất trong tế bào xảy ra nhanh. Quá trình chuyển hóa của VSV sản sinh ra các loại enzym khác nhau và là nguồn thu lớn của các enzym tự nhiên. Protease là một trong 3 nhóm enzym công nghiệp lớn nhất, chiếm khoảng 60% tổng doanh số enzym toàn cầu; hàng trăm protease đã được thương mại hóa và sử dụng trong chất tẩy rửa, chế biến thực phẩm, phụ gia thức ăn chăn nuôi, chế biến da, xử lý chất thải, dược lý và sản xuất thuốc (Sindhu và cs, 2018). Cellulase là enzym quan trọng cả trong công nghiệp và tự nhiên, đóng vai trò chính trong chu trình carbon toàn cầu. Quá trình thủy phân bằng cellulase có thể phục vụ mục đích “kép”: giảm chất thải thực vật, chuyển đổi sinh học thành nhiên liệu và thu hẹp sự phụ thuộc ngày càng tăng vào hóa thạch nhiên liệu, sử dụng trong công nghiệp bột giấy, thực phẩm, rượu vang… Một số VNNS sinh cellulose như Pestalotiopsis sp., Microsphaeropsis sp., Sclerocystis sp., Cephalosporium sp., Penicillium sp., Fusarium oxysporum, Aspergillus sp., Penicillium chrysogenum, Xylaria sp… đã được phân lập từ các cây Acanthus ilicifolius, Zea mays, Sabina chinensis, Taxus chinensis, Keteleeria evelyniana, Pinus massoniana… (Fatima và cs, 2022; Sindhu và cs, 2018). Amylase là enzym thủy phân liên kết alpha của các polysaccharide tạo glucose và maltose; được ứng dụng trong thực phẩm, đồ uống, y tế và được sản xuất tự nhiên bởi nhiều loài nấm khác nhau; trong đó có các loài VNNS P. microspore, A. oryzae và P. chrysogenum trong các cây Rhizophora mucronata, Avicennia ofcinalis, A. marina và Asclepias sinaica (Fatima và cs, 2022; Cripwell và cs, 2021). Lipase là enzym phân hủy chất béo trung tính thành axit béo tự do và glycerol, có ứng dụng lớn trong ngành công nghiệp thực phẩm: làm tăng năng suất chế biến dầu thực vật, tăng hương thơm trong công nghiệp làm bánh và sữa. Nguồn lipase tốt nhất là được khai thác từ nhiều loài nấm như Rhizopus, Mucor, Geotrichum, Pencillium, Aspergillus, Humicola; ngoài ra có các loài VNNS R. oryzae, Cercospora kikuchii, Lasiodiplodia theobromae từ các cây Tithonia diversifolia và Cocos nucifera (Fatima và cs, 2022; Sindhu và cs, 2018). Khả năng sử dụng phosphate không hòa tan trong đất có thể được cải thiện bằng cách sử dụng enzym phosphatase giúp cây trồng sinh trưởng và phát triển tốt hơn; Fitriyana và Ainy (2019) đã phân lập được chủng VNNS từ rễ cây R. mucronate có hoạt tính phosphatase. β-Galactosidase là một exoglycosidase có tác dụng thủy phân liên kết β-glycosid được hình thành giữa galactose và phần hữu cơ của nó; β-galactosidase được sử dụng trong các sản phẩm từ sữa như sữa chua, kem chua và một số loại pho mát được xử lý bằng enzym để phân hủy bất kỳ loại đường lactose nào trước khi tiêu thụ cho con người (Eriana và cs, 2000).
Các kết quả nghiên cứu về các loài VNNS cây H. javanica có khả năng sinh các enzym trên là hầu như chưa có công bố. Các kết quả khả quan này cho thấy đây có thể là nguồn nguyên liệu cho các nghiên cứu sâu hơn để có thể thu được hiệu suất cao các enzym tự nhiên nhằm hướng tới các ứng dụng trong sản xuất nông-công-ngư nghiệp bền vững như ứng dụng trong tạo thức ăn chăn nuôi, xử lý phế phụ phẩm nông-công-ngư nghiệp thành phân bón hữu cơ phục vụ trồng trọt và thay thế phân hóa học cải tạo đất...
3.2. Khả năng kháng VSV gây bệnh của các chủng vi nấm nghiên cứu
Đánh giá khả năng kháng VSV gây bệnh của 9 chủng vi nấm nghiên cứu với 5 chủng VSV kiểm định, bao gồm: 2 chủng vi khuẩn gram dương (B. cereus, S. aureus), 2 chủng vi khuẩn gram âm (E. coli, P. aerigunosa) và 1 chủng nấm men C. albicans. Kết quả thể hiện ở Bảng 2 và Hình 3. cho thấy, 7 chủng nấm (77,77%) có khả năng kháng vi khuẩn B. cereus; 6 chủng (66,66%) kháng vi khuẩn E. coli cũng như kháng nấm men C. albican; 5 chủng (55,55%) kháng vi khuẩn P. aerigunosa; 4 chủng có khả năng kháng S. aureus. Các chủng đều có hoạt tính tiềm năng kháng VSV gây bệnh kiểm định có đường kính vùng ức chế dao động từ 9 ± 0,4 đến 37 ± 1,4 mm. Trong đó, chủng TLC13 có khả năng ức chế cả 5 chủng VSV kiểm định thử nghiệm với hoạt tính gần như cao nhất với 3 loài VSV kiểm định từ 24 ± 1,2 mm (B. cereus), 26 ± 1,1 mm (P. aerigunosa) và 36 ± 1,5 mm (C. albican); 2 chủng TLC10 và TLC19 đều có khả năng kháng 4/5 VSV kiểm định, chủng TLC19 kháng mạnh với B. cereus, P. aerigunosa và C. albican; 4 chủng kháng 3/5 VSV thử nghiệm và chủng chỉ kháng duy nhất 1 loại VSV E. coli là chủng TLC14.
Bảng 2. Khả năng kháng VSV gây bệnh của các chủng nấm nghiên cứu
|
TT |
Chủng vi nấm |
Vùng ức chế sự phát triển của VSV gây bệnh kiểm định (mm) |
||||
|
Vi khuẩn G (+) |
Vi khuẩn G (-) |
Nấm men |
||||
|
B. cereus |
S. aureus |
E. coli |
P. aerigunosa |
C. albican |
||
|
|
TLC9 |
- |
- |
16±0,9 |
20±1,1 |
28±1,4 |
|
|
TLC10 |
15±0,5 |
- |
17±0,4 |
23±0,9 |
17,5±0,3 |
|
|
TLC11 |
13±0,4 |
15±0,6 |
- |
- |
- |
|
|
TLC12 |
17±0,5 |
16±0,7 |
14±0,4 |
- |
- |
|
|
TLC13 |
24±1,2 |
13±0,6 |
16±0,5 |
26±1,1 |
36±1,5 |
|
|
TLC14 |
- |
- |
26±0,9 |
- |
- |
|
|
TLC19 |
22±0,9 |
12±0,3 |
- |
25,5±1,2 |
37±1,4 |
|
|
TLC20 |
18±0,8 |
- |
25±1,2 |
- |
33±1,3 |
|
|
TLC22 |
9±0,4 |
- |
- |
18±0,9 |
28±1,2 |
|
|
ĐC (+) |
18±0,8 |
24±1,1 |
16±0,5 |
23±1,2 |
36±1,3 |
|
|
ĐC (-) |
- |
- |
- |
- |
- |
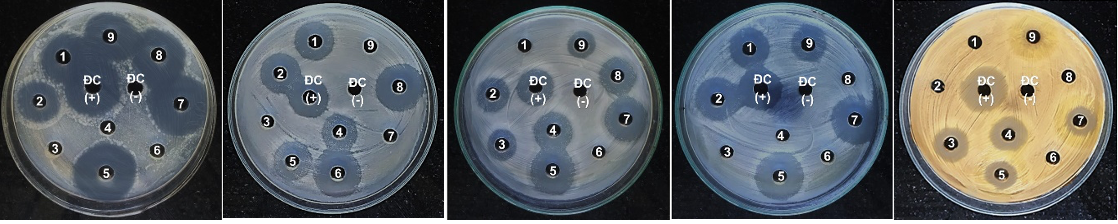
Hình 3. Khả năng kháng VSV gây bệnh kiểm định C. albican, E. coli, B. cereus, P. aerigunosa, S. aureus của các chủng nấm (1. TLC9, 2. TLC10, 3. TLC11, 4. TLC12, 5. TLC13, 6. TLC14, 7. TLC19, 8. TLC20, 9. TLC22, ĐC (-): đối chứng âm, ĐC (+): đối chứng dương)
Từ lâu, các chất kháng sinh có nguồn gốc từ nấm sợi đã được con người biết đến và ứng dụng hiệu quả trong trị bệnh cho người, vật nuôi, cây trồng. Việc tìm kiếm chất kháng sinh mới có nguồn gốc tự nhiên từ các VSV đang được quan tâm. Theo Balick and Cox, năm 1996 trong 119 loại hợp chất hóa học có ít nhất 90 loại có nguồn gốc từ thực vật, đây là các loại thuốc đang được sử dụng ngày càng nhiều ở các quốc gia. Chủng nấm Nigrospora sphaerica URM-6060 phân lập từ lá cây thuốc Indigofera suffruticosa tạo ra hoạt chất sinh học có tiềm năng dược phẩm như tanin thủy phân, alkaloit, dẫn xuất cinnamic có hoạt tính kháng khuẩn với cả vi khuẩn Gram (-) và Gram (+) (Santos và cs, 2015). Jouda và cs (2014) đã phân lập được ba polypetide là penealidins A-C (134-136) có hoạt tính chống lại Acinetobacter sp. và E. coli từ nấm nội sinh Penicillium sp. CAMMC64 phân lập từ lá cây Garcinia nobilis (Clusiaceae) phân bố tại Camơrun. Chiết xuất của các chủng VNNS E. nigrum, F. Tricinctum và Phoma sp. phân lập từ các cây Dendrobium devonianum và D. thyrsiflorum có khả năng kháng lại các vi khuẩn B. subtilis, C. albicans, E. coli và S. aureus; chủng Alternaria sp., Bjerkandera sp., Diaporthe sp., Penicillium sp. và Xylaria sp. phân lập từ cây Schinus terebinthifolius có khả năng kháng lại C. albicans, P. aeruginosa và S. Aureus (Daniel và cs, 2022).
Các nghiên cứu về các loài VNNS kháng VSV gây bệnh ở cây H. javanica chưa có công bố. Các chủng VNNS có phổ kháng rộng và hoạt lực kháng tiềm năng như TLC13, TLC19 và TLC10 có thể sẽ là nguồn nguyên liệu tiềm năng cho việc tìm kiếm và phát hiện các hoạt chất kháng khuẩn mới có thể ứng dụng trong đời sống con người như trong lĩnh vực chăn nuôi, bảo quản chế biến sau thu hoạch…
4. KẾT LUẬN
Nghiên cứu ứng dụng các chủng VNNS thực vật (đặc biệt là VNNS ở các cây thảo dược) trong kinh tế tuần hoàn là một hướng nghiên cứu tiềm năng. Các nghiên cứu về VNNS cây Thạch tùng javanica có khả năng sinh enzym và kháng VSV gây bệnh hiện chưa có công bố. Nghiên cứu này đã sàng lọc và tuyển chọn được một số chủng VNNS cây Thạch tùng javanica Việt Nam có khả năng sinh tổng hợp đa enzym ngoại bào và đa kháng VSV gây bệnh. Trong 6 emzym thử nghiệm, chủng TLC11 cho khả năng sinh 5/6 loại ngoại trừ enzym amylase; tiếp theo là chủng TLC9 sinh 4/5 loại enzym; 4 chủng sinh 3 loại enzym là TLC10, TLC19, TLC20 và TLC22. Protease và lipase là 2 hoạt tính được thể hiện mạnh nhất ở các chủng sinh hoạt tính, có đường kính vòng thủy phân từ 22÷34 mm (protease) và 20÷28 mm (lipase). Trong 5 loại VSV gây bệnh kiểm định, chủng TLC13 ức chế cả 5 chủng VSV kiểm định với hoạt tính gần như cao nhất với 3 loài B. cereus (24±1,2 mm), P. aerigunosa (26±1,1 mm) và C. albican (36±1,5 mm); 2 chủng TLC10 và TLC19 kháng 4/5 VSV kiểm định ngoại trừ S. aureus (TLC10) và E. coli (TLC19); 4 chủng kháng 3/5 VSV thử nghiệm. Các chủng nghiên cứu có khả năng sinh các enzym ngoại bào với hoạt tính mạnh, có phổ kháng VSV gây bệnh tương đối rộng với hoạt lực ức chế tiềm năng là TLC9, TLC11, TLC13, TLC10, TLC19. Các chủng này sẽ là nguồn nguyên liệu tiềm năng ứng dụng trong các lĩnh vực của ngành nông - công - ngư nghiệp - dược phẩm bền vững. Tuy nhiên, cần tiếp tục có các nghiên cứu sâu hơn như xác định các hoạt chất kháng khuẩn, hoạt lực của các chất kháng khuẩn và enzym ngoại bào, các điều kiện lên men tăng năng suất và thu nhận enzym/chất kháng khuẩn cũng như các thử nghiệm ứng dụng chúng trong thực tiễn, từ đó đưa ra giải pháp ứng dụng cụ thể cho từng ngành nghề như chăn nuôi, trồng trọt, bảo quản chế biến sau thu hoạch, công nghiệp thực phẩm và xử lý môi trường.
Lời cảm ơn: Nghiên cứu này được hỗ trợ kinh phí từ đề tài hợp phần trọng điểm cấp Viện Hàn lâm Khoa học và Công nghệ Việt Nam, mã số TĐCNSH.04/20-22.
Trịnh Thị Thu Hà1, Phạm Thanh Hà1, Hoàng Thị Yến1, Lê Thị Minh Thành1*
1Viện Công nghệ sinh học, Viện Hàn lâm Khoa học và Công nghệ Việt Nam
(Nguồn: Bài đăng trên Tạp chí Môi trường, số Chuyên đề Tiếng Việt I/2024)
TÀI LIỆU THAM KHẢO
1. Balick M and Cox PA (1996). Plants, People, and Culture: A Scientific American Library Volume. Freeman, New York. 228 pages. ISBN: 0-7167-5061-9.
2. Cripwell RA, Heber WZ, Viljoen-Bloom M (2021). Fungal Biotechnology: Fungal Amylases and Their Applications. Volume 2, p. 326-336.
3. Daniel JC, Enzo AP, Simon EM, Bita Z (2022). Exploring the promise of endophytic fungi: a review of novel antimicrobial compounds. Microorganisms 10, 1990 (1-22). https://doi.org/10.3390/microorganisms10101990.
4. Fatima B, Anu G, Vasundhara M, Sudhakara RM (2022). Endophytic fungi: a potential source of industrial enzyme producers. Biotech 12 (86): 1-17. https://doi.org/10.1007/s13205-022-03145-y.
5. Fitriyana H and Ainy EQ (2019). Cultivable Endophytic Fungi Producing Phosphatase of Rhizophora mucronate. J. Phys.: Conference Series, DOI 10.1088/1742-6596/1594/1/012002.
6. Inamuddin, Mohd IA, Ram P (2022). Application of Microbes in Environmental and Microbial Biotechnology. Publisher: Springer Singapore, Edition No 1, p. IX-736.
7. Jouda JB, Kusari S, Lamshöft M, Mouafo TF, Douala MC, Wandji J, Spiteller M (2014). Penialidins A-C with strong antibacterial activities from Penicillium sp., an endophytic fungus harboring leaves of Garcinia nobilis. Fitoterapia. 98:209-214. doi: 10.1016/j.fitote.2014.08.011.
8. Lu Z, Ma Y, Xiao L, Yang H and Zhu D (2021). Diversity of Endophytic Fungi in Huperzia serrata and Their Acetylcholinesterase Inhibitory Activity. Sustainability 13, 12073. https://doi.org/10.3390/su132112073.
9. Quyết định số 882/QĐ-TTg ngày 22/7/2022 của Thủ tướng Chính phủ phê duyệt Kế hoạch hành động quốc gia về tăng trưởng xanh giai đoạn 2021-2030.
10. Quyết định số 687/QĐ-TTg ngày 7/6/2022 của Thủ tướng Chính phủ phê duyệt Đề án Phát triển kinh tế tuần hoàn ở Việt Nam.
11. Santos IP, Silva LCN, Silva MV, Araújo JM, Cavalcanti MS, Lima VLM (2015). Antibacterial activity of endophytic fungi from leaves of Indigofera suffruticosa Miller (Fabaceae). Front. Microbiol. 6:350. doi: 10.3389/fmicb.2015.00350.
12. Sindhu R, Binod P, Sabeela BU, Amith A, Anil KM, Aravind M, Sharrel R, Ashok P (2018). Applications of Microbial Enzyms in Food Industry. Food Technol Biotechnol. 56(1): 16-30.
13. Sun BY, 2015. Lycopodiaceae In: Park CW, editors. Flora of Korea, vol. 1 Incheon, South Korea: National Institute of Biological Resources; p. 13-16.
