28/05/2025
Tóm tắt:
Gò đồi ngầm Cô Tô - Thanh Lân có những đặc thù sinh thái riêng biệt, nguồn lợi sinh vật đáy đa dạng và là nơi khai thác thủy sản của nhiều ngư dân địa phương, tuy nhiên, đến nay chưa có nghiên cứu nào cụ thể ở gò đồi ngầm này để phục vụ quản lý nguồn lợi hiệu quả. Nghiên cứu được thực hiện trong hai đợt khảo sát thực địa bằng phương pháp lặn SCUBA (là hình thức lặn biển sử dụng thiết bị thở độc lập dưới nước, cho phép người lặn thở tự do và khám phá thế giới dưới biển ở độ sâu lớn hơn và trong thời gian dài hơn so với lặn ống thở), thu mẫu thành phần loài và sinh thái, định danh loài và phân tích phân bố sinh thái thực hiện vào năm 2023 và 2024. Kết quả phân tích cho thấy khu vực này có ba dạng nền đáy đặc trưng gồm: dạng nền đáy rạn san hô (ở khu vực đỉnh gò với độ sâu 12 - 15 m); dạng nền đáy đá tảng và cụm san hô (ở khu vực sườn gò, có độ sâu 18 - 22 m); dạng nền đáy cát bùn (ở chân gò với độ sâu từ 25 - 35 m). Tổng số 95 loài động vật đáy thuộc ba nhóm chính là Mollusca, Arthropoda và Echinodermata đã được ghi nhận lần đầu tiên ở gò đồi ngầm Cô Tô - Thanh Lân. Sự phân bố của một số nhóm loài đặc trưng theo dải độ sâu và dạng nền đáy như loài Mauritia arabica tập trung ở vùng đá rạn nông (14 - 20 m), trong khi Atrina vexillum, Pteria penguin và Colochirus quadrangularis hiện diện chủ yếu ở tầng sâu (>25 m). Nghiên cứu cũng ghi nhận có một số loài có giá trị kinh tế như Atrina vexillum, Pteria peasei và Mauritia arabica. Kết quả nghiên cứu này là cơ sở khoa học cho việc đề xuất các giải pháp bảo tồn và quản lý khai thác hợp lý nhằm bảo vệ nguồn lợi sinh vật đáy và hướng tới phát triển kinh tế biển bền vững tại các gò đồi ngầm trong tương lai.
Từ khóa: Cô Tô - Thanh Lân, ĐDSH, động vật đáy, phân bố sinh thái.
Ngày nhận bài: 2/4/2025; Ngày sửa chữa: 28/4/2025. Ngày duyệt đăng: 27/5/2025.
SPECIES DIVERSITY AND ECOLOGICAL DISTRIBUTION OF ZOOBENTHOS AT THE CO TO - THANH LAN SEAMOUNT, QUANG NINH, VIETNAM
Abstract:
The Co To - Thanh Lan seamount possesses unique ecological characteristics, a diverse benthic fauna, and important fishing grounds for many local fishermen. However, to date, no specific studies have been conducted on this seamount to support effective resource management. This study was carried out through two field surveys using SCUBA diving methods to collect data on species composition, ecology, species identification, and ecological distribution in 2023 and 2024. The analysis results show that the area has three distinct substrate types: Coral reef substrate (at the top of seamount, 12–15 meters deep); Rocky and coral cluster substrate (on the seamount slopes, 18–22 meters deep); Sandy-muddy substrate (at the base of seamount, 25–35 meters deep). A total of 95 benthic animal species from three main groups (Mollusca, Arthropoda, and Echinodermata) were recorded for the first time in the Co To – Thanh Lan seamount area. The distribution of some species was associated with depth ranges and substrate types, such as Mauritia arabica, which was concentrated in shallow rocky reef areas (14–20 m), while Atrina vexillum, Pteria penguin, and Colochirus quadrangularis were primarily found at greater depths (>25 m). The study also recorded several economically valuable species, including Atrina vexillum, Pteria peasei, and Mauritia arabica. These findings provide a scientific foundation for proposing conservation measures and sustainable exploitation strategies to protect benthic animal resources and promote sustainable marine economic development at seamounts in the future.
Keywords: Biodiversity, Co To - Thanh Lan, Distribution, Zoobenthos.
JEL Classifications: O13, P48, Q57.
1. Mở đầu
Các hệ sinh thái (HST) rạn ngầm và gò đồi ngầm được công nhận trên toàn cầu là các điểm nóng đa dạng sinh học (ĐDSH), đóng vai trò thiết yếu trong việc duy trì các quần thể thủy sản và cung cấp các dịch vụ HST (Roberts, 2002)(Harris, 2014). Những khu vực này thường là nơi sinh sống của nhiều loài động vật đáy như động vật giáp xác, thân mềm, san hô và hải miên, góp phần vào chuỗi thức ăn và cấu trúc sinh thái biển (Pitcher, 2016). Theo các tiêu chí về sinh thái học của IUCN (2012) đã nhấn mạnh các đặc điểm đặc trưng về môi trường sinh thái các gò đồi ngầm, gồm: (a) Là khu vực có nguồn dinh dưỡng phong phú; (b) là nơi có điều kiện thủy văn đặc thù với các dòng xoáy mạnh và nước trồi cục bộ; (c) là nơi cung cấp các sinh cảnh hết sức phong phú do có hệ thống các hang ngầm hết sức phức tạp; (d) là bãi đẻ của các loài cá, nơi kiếm ăn của các loài động vật biển; (e) là khu vực tiềm tàng ẩn chứa các loài sinh vật biển đặc hữu; (f) là nơi có tiềm năng đánh bắt thủy sản do phần lớn nằm ở các khu vực có ngư trường trọng điểm. Tuy nhiên, trong những thập kỷ gần đây, dưới sức ép do tác động tiêu cực của hoạt động con người như kéo lưới đáy trong khai thác thủy sản đã trở thành mối đe dọa đến ĐDSH và khả năng phục hồi của HST này (Rogers, 2019).
Ở Việt Nam, trong thời gian qua, đã có nhiều nghiên cứu về các HST biển bao gồm: Rạn san hô; rừng ngập mặn; thảm cỏ biển; cửa sông và bãi bồi ven biển; vùng triều; đầm phá. Trong khi đó HST bãi cạn, gò đồi ngầm có tầm quan trọng lớn trong việc duy trì ĐDSH biển, duy trì tính sinh học đặc hữu ở vùng biển Việt Nam nhưng gần như chưa có những thông tin, khám phá về ĐDSH trong HST này. Do đó, công tác điều tra ĐDSH và nguồn lợi hải sản tại các bãi cạn, gò đồi ngầm ở vùng biển Việt Nam là rất cần thiết, nhằm định hướng khai thác tiềm năng, lợi thế của gò đồi ngầm trong phát triển kinh tế biển, đặc biệt là kinh tế nghề cá. Trong Vịnh Bắc Bộ, vùng biển Cô Tô - Thanh Lân được ghi nhận có một số gò đồi ngầm có thành phần loài động vật đáy đa dạng, rạn san hô phát triển và đang chịu sức ép khai thác từ người dân địa phương. Tuy nhiên, đến nay, chưa có những nghiên cứu công bố cụ thể về hình thái rạn, kiểu nền đáy, thành phần loài động vật đáy và đặc trưng phân bố của chúng.
Trong bối cảnh như vậy, khu vực gò đồi ngầm Cô Tô - Thanh Lân (Quảng Ninh), được lựa chọn nghiên cứu nhằm đánh giá hiện trạng ĐDSH phục vụ quản lý và bảo tồn tài nguyên biển. Đây là một trong những nỗ lực nghiên cứu đầu tiên tại Việt Nam về khu HST gò đồi ngầm vùng biển phía Bắc. Một số kết quả nghiên cứu tại gò đồi ngầm Cô Tô - Thanh Lân được trình bày trong báo cáo này bao gồm: (1) Thành phần loài của nhóm động vật đáy, (2) Đặc điểm phân bố sinh thái và (3) Mối liên quan giữa các đặc điểm nền đáy với sự phân bố của khu hệ động vật đáy. Kết quả nghiên cứu này là cơ sở khoa học để đề xuất các biện pháp quản lý và bảo tồn hiệu quả, đồng thời đóng góp vào chiến lược phát triển kinh tế biển bền vững của vùng biển Quảng Ninh nói riêng và vùng biển Việt Nam nói chung.
2. THỜI GIAN VÀ PHƯƠNG PHÁP NGHIÊN CỨU
2.1. Địa điểm và thời gian khảo sát
Nghiên cứu được tiến hành tại khu vực gò đồi ngầm Cô Tô - Thanh Lân, thuộc vùng biển phía Bắc Việt Nam. Các đợt khảo sát hiện trường được thực hiện vào tháng 11/2023 và tháng 5/2024, đại diện cho hai mùa chính (mùa khô và mùa mưa) nhằm ghi nhận sự biến đổi theo mùa trong khu hệ động vật đáy. Vị trí các điểm khảo sát và phân bố dải độ sâu được xác định bằng máy định vị GPS Garmin (Garmin Ltd., USA), theo phương pháp được mô tả bởi (English, 1997), đảm bảo độ chính xác tọa độ địa lý cho các điểm thu mẫu. Tổng số mặt cắt khảo sát là 12 (Hình 1).
2.2. Đối tượng nghiên cứu
Nghiên cứu tập trung vào ba nhóm khu hệ động vật đáy chính: Chân khớp (Arthropoda), thân mềm (Mollusca) và da gai (Echinodermata). Các nhóm này được chọn do vai trò sinh thái quan trọng của chúng trong HST gò đồi ngầm, bao gồm đóng góp vào ĐDSH, chuỗi thức ăn và cấu trúc môi trường sống (Pitcher, 2016).
2.3. Phương pháp nghiên cứu
Phương pháp thu mẫu và đặc điểm phân bố sinh thái
Mẫu động vật đáy được thu thập tại nền đáy của khu vực gò đồi ngầm bằng phương pháp lặn SCUBA, sử dụng kỹ thuật dây mặt cắt (transect) với phạm vi khảo sát 2,5 m mỗi bên dây, theo phương pháp thu mẫu định lượng và định tính (English, 1997). Tại mỗi điểm khảo sát, tất cả các loài thuộc ba nhóm động vật đáy (Chân khớp, thân mềm, da gai) được thu thập dọc theo dây mặt cắt và vùng lân cận trong khu vực khảo sát. Vị trí từng điểm thu mẫu được ghi lại bằng tọa độ GPS để đảm bảo khả năng tái định vị và phân tích không gian.
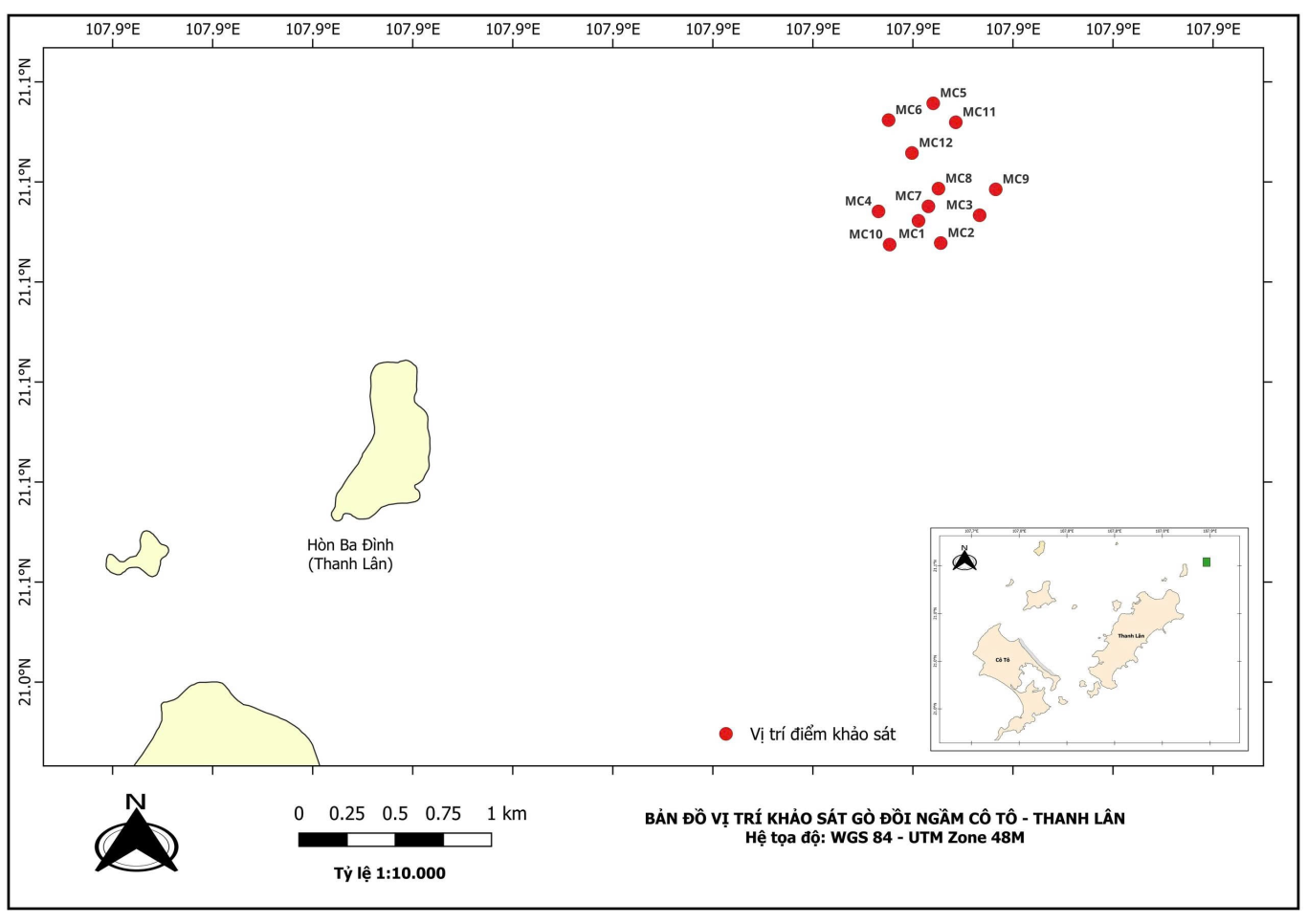
Hình 1. Khu vực khảo sát gò đồi ngầm tại vùng biển Cô Tô - Thanh Lân (Quảng Ninh)
Cố định và lưu trữ mẫu
Sau khi thu thập, các mẫu được rửa sạch bằng nước biển tại hiện trường để loại bỏ trầm tích và tạp chất. Mẫu vật được cố định ngay lập tức trong dung dịch ethanol 70% để bảo quản giữ nguyên vẹn đặc điểm hình thái. Mỗi mẫu được gắn nhãn riêng, ghi rõ thông tin về vị trí thu mẫu (tọa độ GPS), ngày thu mẫu và đặc điểm môi trường (độ sâu, loại nền đáy). Các mẫu sau đó được vận chuyển về phòng thí nghiệm tại Viện Nghiên cứu Hải sản (RIMF, Việt Nam) để phân tích chi tiết.
Định danh loài sinh vật và xử lý số liệu
Sử dụng phần mềm QGIS (phiên bản 3.22) để xây dựng bản đồ phân bố địa hình của gò đồi ngầm và phân tích sự phân bố không gian của các loài động vật đáy. Bản đồ được tạo dựa trên dữ liệu độ sâu và đặc điểm trầm tích từ các khảo sát hiện trường, kết hợp với dữ liệu viễn thám (nếu có) để mô tả đặc điểm địa hình và môi trường sống.
Tên khoa học của các loài thuộc ba nhóm động vật đáy được xác định dựa trên phương pháp hình thái, bằng các tài liệu phân loại phổ biến (Kent E. Carpenter, 1998) (Carpenter, 1998a, 1998b), (Okutani, 2000) và (Terrence, 1996).
3. KẾT QUẢ NGHIÊN CỨU
3.1. Đặc điểm địa hình gò đồi ngầm Thanh Lân - Cô Tô
Kết quả khảo sát tại khu vực bãi cạn và gò đồi ngầm Thanh Lân - Cô Tô cho thấy khu vực này có một đỉnh gò duy nhất với độ sâu dao động từ 9 m đến 14 m tại vị trí nông nhất. Tổng diện tích toàn bộ khu vực gò đồi ngầm được xác định trong khoảng 1,0 đến 1,5 km², với chiều rộng dao động từ 500 m đến 1.000 m và chiều dài từ 1.500 m đến 2.000 m, Bản đồ độ sâu (Hình 2) cho thấy sự phân bố độ sâu không đồng đều, với khu vực trung tâm gò có độ sâu nông nhất (9 - 14 m) và độ sâu tăng dần về phía chân gò, đạt từ 35 m đến 45 m. Nền đáy tại khu vực khảo sát khá đa dạng, với đá pha cát ở đỉnh rạn và chuyển sang cát bùn ở chân rạn, kèm thưa thớt những khối đá tảng lớn, điều này thể hiện được sự đa dạng của hình thái rạn ngầm tại vùng biển Cô Tô - Thanh Lân.
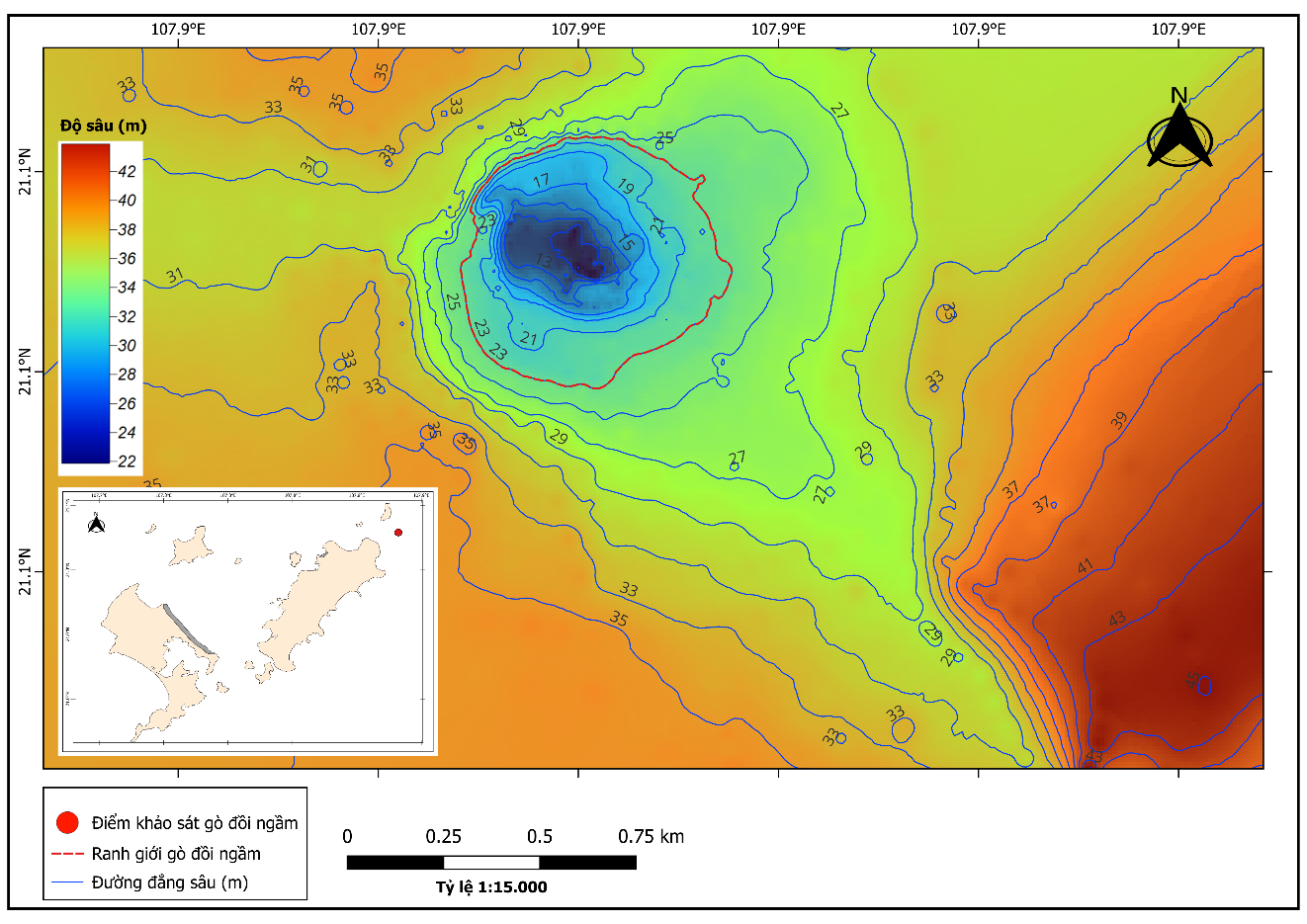
Hình 2. Bản đồ hình thái gò đồi ngầm theo độ sâu (m) tại vùng biển Cô Tô - Thanh Lân
Dựa trên các tài liệu phân loại dạng gò đồi ngầm trên thế giới, gò đồi ngầm Cô Tô - Thanh Lân có thể được phân loại như sau: Hình thái: Kiểu hình nón bất quy tắc (Wessel, 2010), do có một đỉnh gò duy nhất nhưng hình thái nền đáy không đồng đều. Gò đồi ngầm có dạng dốc thẳng đứng ở hướng Tây Bắc, và có dạng sườn thoải kéo dài hướng Đông Nam (Hình 3). Độ sâu: Thuộc nhóm gò nông đến trung bình (Yesson et al., 2011), với độ sâu đỉnh gò từ 9 - 14 m và chân gò từ 35 - 45 m. Đặc điểm nền đáy: Bao gồm nền đá gốc (đá lẫn cát, đá tảng nhỏ xen bùn mịn) và nền trầm tích cát bùn mịn ở vùng chân gò (Harris, 2014).
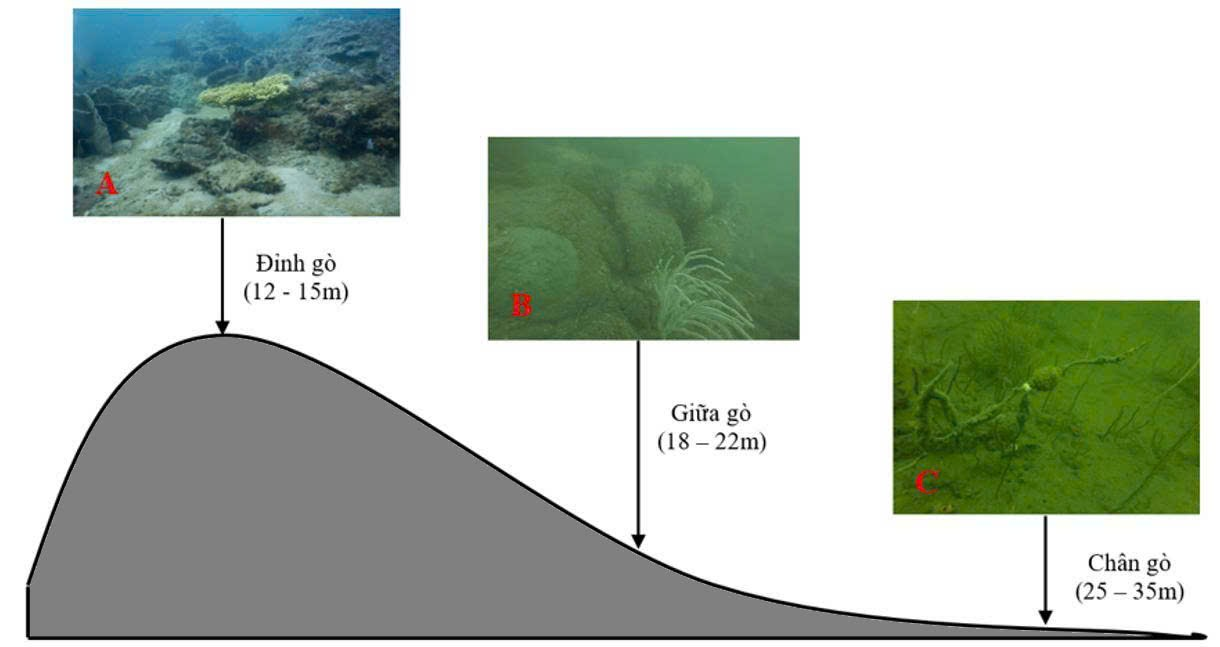
Hình 3. Mặt cắt ngang và các dạng nền đáy của gò đồi ngầm Cô Tô - Thanh Lân (Quảng Ninh). (A) là dạng nền đáy rạn san hô ở khu vực đỉnh gò đồi ngầm (12 - 15m); (B) là dạng nền đáy đá tảng và cụm san hô ở khu vực bên sườn gò đồi ngầm (18 - 22m); (C) là dạng nền đáy cát bùn ở chân gò đồi ngầm (25 - 25m)
Dựa trên đặc điểm hình thái và độ sâu, khu vực gò đồi ngầm Cô Tô - Thanh Lân được phân chia thành ba dạng hình thái rạn tương đối khác biệt gồm: (A) là dạng nền đáy rạn san hô ở khu vực đỉnh gò đồi ngầm (sâu 12 - 15m); (B) là dạng nền đáy đá tảng và cụm san hô ở khu vực bên sườn gò đồi ngầm (sâu 18 - 22m); (C) là dạng nền đáy cát bùn ở chân gò đồi ngầm (sâu 25 - 25m). Đặc điểm nền đáy có cả san hô, đá tảng và cát bùn này thuận lợi cho sự phát triển của các loài sinh vật đáy.
Tại khu vực chân gò đồi ngầm Thanh Lân, nằm ở hướng Đông Nam của gò đồi ngầm Thanh Lân, với nền đáy đặc trưng bởi đá tảng nhỏ mật độ cao xen lẫn bùn mịn (Hình 3C). Thành phần đá tảng nhỏ chiếm ưu thế (khoảng 60 - 70%), trong khi bùn mịn phân bố xen kẽ, tạo nên một môi trường sống đa dạng. Độ sâu tại khu vực này dao động từ 25 m đến 35 m. Đặc điểm nền đáy này thuận lợi cho các loài sinh vật sống trong nền bùn, chẳng hạn như các loài thân mềm (Mollusca) và da gai (Echinodermata), nhờ vào sự tích tụ chất hữu cơ trong lớp bùn mịn.
Ngoài ra, khu vực bao quanh phần chân gò có độ sâu từ 35 m đến 45 m, với nền đáy chủ yếu là cát bùn mịn, được xem là vùng chuyển tiếp quan trọng trong HST gò đồi ngầm (Hình 3). Lớp cát bùn mịn tại đây đóng vai trò như một bể chứa phù sa, hỗ trợ quá trình tích tụ chất dinh dưỡng và tạo môi trường sống phù hợp cho nhiều loài động vật đáy, bao gồm các loài giáp xác (Arthropoda) và thân mềm (Mollusca). Khu vực này cũng đóng vai trò như một vùng đệm, kết nối giữa gò đồi ngầm và các HST biển sâu hơn, góp phần duy trì ĐDSH và chuỗi thức ăn trong khu vực.
3.2. Đa dạng thành phần loài
Kết quả nghiên cứu thành phần loài động vật đáy thu được ở khu vực gò đồi ngầm tại vùng biển Cô Tô - Thanh Lân, Quảng Ninh đã xác định được 95 loài, thuộc 71 giống, 49 họ, 26 bộ, 10 lớp thuộc 3 ngành chính là chân khớp (Arthropoda), da gai (Echinodermata) và thân mềm (Mollusca). Trong đó, ngành chân khớp (Arthropoda) chiếm ưu thế với tỷ lệ cao nhất là 57,89%. Sự phân bố ưu thế của ngành này tại đây có thể liên quan đến sự thích nghi của nhóm này với nền đáy đa dạng (đá lẫn cát, đá tảng xen bùn mịn) tại khu vực phần cuối của gò đồi ngầm. Nhóm ngành da gai (Echinodermata) chiếm 22,11%, đứng thứ hai về tỷ lệ. Chúng thường bao gồm các loài như sao biển, cầu gai và thường phân bố nhiều ở các khu vực nằm ở phần cuối của gò đồi ngầm Thanh Lân, tại nơi có nền đáy là đá tảng nhỏ xen bùn mịn thường với độ sâu 25 - 35 m. Nhóm ngành Mollusca (Thân mềm) chiếm 20,00%, thấp nhất trong ba nhóm. Nhóm này bao gồm các loài như ngao, sò, ốc, có thể bị hạn chế bởi điều kiện môi trường hoặc cạnh tranh sinh học với nhóm Chân khớp.
Sự chiếm ưu thế của nhóm Arthropoda (gần 58%) cho thấy khu vực gò đồi ngầm này có thể là môi trường sống lý tưởng cho các loài giáp xác (như tôm, cua), nhờ vào nền đáy đa dạng và sự tích tụ chất hữu cơ ở vùng chân gò (độ sâu 35 - 45 m). Tỷ lệ tương đối cân bằng giữa Echinodermata và Mollusca (22,11% và 20,00%) cho thấy sự ĐDSH trong khu vực, nhưng cũng có thể phản ánh sự cạnh tranh giữa các nhóm này. Hai họ Muricidae và Portunidae có số lượng loài đa dạng nhất có 8 loài (chiếm 11,59% tổng số loài), ba họ Pinnidae, Margaritidae và Temnopleuridae có 4 loài (chiếm 5,80% tổng số loài). Các họ còn lại ghi nhận từ 1 - 3 loài/họ.Trong số các loài đã ghi nhận, nhóm động vật thân mềm Mollusca ghi nhận số lượng loài cao nhất (gồm 14 bộ, 26 họ, 39 giống, 54 loài, chiếm 56,84%), tiếp theo là nhóm động vật chân khớp Arthropoda có (5 bộ, 12 họ, 17 giống, 22 loài, chiếm 23,16%), nhóm động vật da gai Echinodermata có (7 bộ, 11 họ, 15 giống, 19 loài, chiếm 20%).
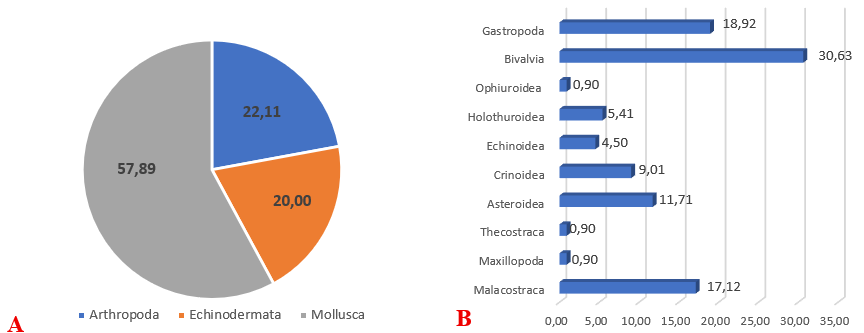
Hình 4. Tỷ lệ % thành phần loài của nhóm động vật đáy phân bố tại gò đồi ngầm Cô Tô - Thanh Lân. (A) Tỷ lệ % các nhóm ngành sinh vật đáy; (B) Tỷ lệ % các lớp động vật đáy
Khi xem xét sự phân bố của các lớp động vật đáy tại vùng gò đồi ngầm Cô Tô - Thanh Lân cho thấy, Lớp Chân bụng (Gastropoda) chiếm 30,63%. Đây là lớp chiếm tỷ lệ cao nhất. Lớp Giáp xác (Malacostraca) chiếm 17,12%. Thành phần thuộc lớp này phản ánh sự hiện diện đáng kể của các loài tôm, cua, phù hợp với tỷ lệ cao của nhóm Arthropoda (Hình 4). Điều này có thể liên quan đến sự đa dạng nền đáy và nguồn thức ăn phong phú. Các lớp có tỷ lệ thấp hơn gồm: Lớp hải sâm (Holothuroidea) chiếm 5,41%, thường gặp ở nền đáy bùn mịn ở vùng chân gò), Lớp Cầu gai (Echinoidea) với 4,50%, thường thấy ở nền đáy cứng. Lớp Sao biển (Asteroidae) chỉ có 11,71%, thường bắt gặp tại các khu vực nền đáy đá lẫn cát ở khu vực đỉnh gò và giữa gò đồi ngầm, ở độ sâu 12–22 m. Các lớp khác như Lớp Hai mảnh vỏ (Bivalvia), Lớp Đuôi rắn (Ophiuroidea), Lớp Huệ biển (Crinoidea), Lớp Thecostraca, mỗi lớp chiếm 0,90%, cho thấy sự hiện diện rất hạn chế, có thể do cạnh tranh sinh học hoặc điều kiện môi trường không thuận lợi cho các lớp này tại gò đồi ngầm Cô Tô - Thanh Lân.
Năm 2023, nhóm nghiên cứu khảo sát tại khu vực gò đồi ngầm Cô Tô - Thanh Lân (Quảng Ninh) đã ghi nhận được 33 loài động vật đáy, trong đó nhóm loài động vật thân mềm (Mollusca) chiếm ưu thế với 21 loài, tiếp theo là nhóm loài động vật chân khớp (Arthropoda) với 5 loài và nhóm loài động vật da gai (Echinodermata) với 7 loài. Năm 2024, số lượng loài ghi nhận tăng đáng kể, đạt tổng cộng 74 loài, gồm 45 loài động vật thân mềm, 16 loài động vật chân khớp và 13 loài động vật da gai. Sự gia tăng này phần lớn do khác biệt về thời điểm khảo sát: năm 2023 được thực hiện vào tháng 11 (cuối thu – đầu đông), trong khi năm 2024 tiến hành vào tháng 5 (cuối xuân – đầu hè), là giai đoạn nhiều loài sinh vật đáy bước vào thời kỳ hoạt động mạnh và sinh sản. Kết quả này phù hợp với nhận định trong nghiên cứu của (Nguyễn Văn Quân, 2012), trong đó họ đã khảo sát đa dạng sinh vật đáy tại khu vực Cô Tô và Thanh Lân. Nghiên cứu của Nguyễn Văn Quân chỉ ra rằng vào mùa hè, sự phong phú loài và mật độ sinh vật đáy tăng cao hơn đáng kể so với các mùa thu và đông, do điều kiện nhiệt độ và dinh dưỡng thuận lợi thúc đẩy sự phát triển và sinh sản của nhiều loài. Ngoài ra, các loài thân mềm như các loài trai, sò và ốc có xu hướng hoạt động và sinh trưởng mạnh mẽ hơn vào mùa xuân - hè, góp phần nâng cao ĐDSH trong thời điểm này.
Tương tự, nghiên cứu của (Nguyễn Văn Khương, 2015) tại vùng ven biển Quảng Ninh cũng cho thấy sự biến động rõ rệt về thành phần loài và số lượng sinh vật đáy theo mùa vụ. Họ nhận thấy ĐDSH sinh vật đáy đạt mức cao nhất vào mùa xuân – hè, đặc biệt là nhóm thân mềm và chân khớp, trong khi vào mùa thu – đông thì số lượng và sự đa dạng giảm đi đáng kể do điều kiện môi trường như nhiệt độ nước biển và nguồn thức ăn thay đổi. Kết quả này minh chứng rõ ràng rằng yếu tố mùa vụ là nhân tố quyết định ảnh hưởng đến sự biến động về cấu trúc cộng đồng và phân bố sinh vật đáy ở khu vực gò đồi ngầm Cô Tô - Thanh Lân.
3.3. Đặc điểm phân bố
Từ các kết quả khảo sát năm 2023 - 2024, khi phân tích mật độ phân bố của 6 loài động vật đáy ưu thế và đặc hữu tại khu vực gò đồi ngầm Cô Tô - Thanh Lân theo ba dải độ sâu cho thấy xu hướng gia tăng rõ rệt về tỷ lệ hiện diện tại khu vực giữa và chân rạn. Tại khu vực đỉnh rạn (14 - 20 m) với nền đáy đá rạn, chỉ có 4/6 loài được ghi nhận, trong đó Mauritia arabica đạt tỷ lệ cao nhất (60%), còn lại đều có tỷ lệ phân bố rất thấp (0 - 16,7%), hai loài Colochirus quadrangularis và Pteria penguin không băt gặp tại khu vực này. Ở khu vực giữa rạn (20 - 25 m), nơi nền đáy chuyển sang cát bùn - đá tảng, tất cả các loài đều bắt gặp với mật độ từ 14,3% đến 57,1%; nổi bật là Pteria penguin (57,1%), Charybdis japonica và Atrina vexillum (33,3%). Tại khu vực chân rạn (>25 m), mật độ phân bố tăng rõ rệt với Colochirus quadrangularis đạt 71,4%, Pteria peasei 55,6%, Atrina vexillum và Charybdis japonica đều đạt 50% (Bảng 1).
Bảng 1. Phân bố mật độ nhóm loài động vật đáy ưu thế và đặc trưng theo dải độ sâu và nền đáy tại khu vực gò đồi ngầm Cô Tô - Thanh Lân (Quảng Ninh)
|
Độ sâu (m) |
Dạng nền đáy ưu thế |
Loài ưu thế và đặc trưng cho vùng gò đồi ngầm |
|||||
|
Atrina vexillum |
Charybdis japonica |
Mauritia arabica |
Colochirus quadrangularis |
Pteria peasei |
Pteria penguin |
||
|
14 - 20m |
Rạn san hô |
16,7% |
16,7% |
60,0% |
0,0% |
16,7% |
0,0% |
|
20 - 25m |
Đá tảng và cụm san hô |
33,3% |
33,3% |
14,3% |
28,6% |
27,8% |
57,1% |
|
> 25m |
Cát bùn |
50,0% |
50,0% |
25,7% |
71,4% |
55,6% |
42,9% |

Hình 5. Một số loài động vật đáy ưu thế, đặc trưng tại khu vực gò đồi ngầm Cô Tô - Thanh Lân. (A) Atrina vexillum; (B) Charybdis japonica; (C) Mauritia arabica; (D) Colochirus quadrangularis; (E) Pteria peasei; (F) Pteria penguin
Nhìn chung, sự khác biệt trong tỷ lệ bắt gặp và hiện diện của 6 loài tiêu biểu theo các tầng sâu tại gò đồi ngầm Cô Tô - Thanh Lân phản ánh rõ nét tính đặc trưng sinh thái của từng loài và vai trò của điều kiện nền đáy - độ sâu trong việc định hình quần xã sinh vật đáy. Kết quả này là cơ sở quan trọng cho việc quy hoạch bảo tồn sinh cảnh cùng các loài đặc trưng và đánh giá tiềm năng khai thác hợp lý theo từng dạng sinh thái biển.
3.4. Thảo luận
Kết quả khảo sát tại khu vực gò đồi ngầm Cô Tô - Thanh Lân cho thấy nền đáy và độ sâu ảnh hưởng rõ rệt đến cấu trúc quần xã động vật đáy. Ở vùng rạn san hô (14 - 16 m), cá thể có kích thước trung bình và lớn chiếm ưu thế, phản ánh điều kiện sinh cảnh thuận lợi với nhiều nơi trú ẩn và nguồn thức ăn phong phú. Tầng trung gian (20 - 25 m), nền đáy chủ yếu là cụm san hô xen đá tảng, tập trung nhiều cá thể nhỏ hơn, có thể do đây là khu vực cư trú của cá thể non hoặc loài kích thước nhỏ. Ở tầng sâu hơn (26 - 35 m), sự phân bố kích thước cá thể đồng đều hơn, phản ánh môi trường ổn định và ít chịu tác động khai thác. Mô hình này phù hợp với kết quả nghiên cứu của (Bát, 2022) và (Tuấn, 2019), nhấn mạnh vai trò quan trọng của nền đáy và độ sâu trong hình thành cấu trúc quần xã sinh vật đáy.
Kết quả nghiên cứu về khu vực gò đồi ngầm Cô Tô - Thanh Lân đã ghi nhận 95 loài động vật đáy, mức độ ĐDSH tương đối cao. Kết quả này khá tương đồng với nghiên cứu của (Khương, 2005) tại vùng rạn san hô ven đảo Cô Tô với với 100 loài động vật đáy được ghi nhận. Ngoài ra, sự biến động theo mùa vụ rất rõ nét, với 33 loài được ghi nhận vào tháng 11/2023 và 74 loài vào tháng 5/2024, phù hợp với xu hướng phát triển sinh vật đáy trong mùa xuân – hè như kết luận của (Nguyễn Văn Quân, 2012).
Phân tích mật độ phân bố theo độ sâu và nền đáy cho thấy 6 loài đặc trưng gồm Atrina vexillum, Charybdis japonica, Colochirus quadrangularis, Mauritia arabica, Pteria peasei và Pteria penguin có xu hướng gia tăng tỷ lệ bắt gặp ở tầng sâu (>25 m) với nền đáy cát bùn xen lẫn đá. Tại đỉnh gò đồi gầm (14 - 20 m, rạn san hô), mật độ ghi nhận nhìn chung thấp, ngoại trừ Mauritia arabica chiếm ưu thế, cho thấy khả năng thích nghi tốt với môi trường rạn nông hoặc có thể bị ảnh hưởng bởi hoạt động khai thác. Sang tầng sườn gò đồi ngầm (20 - 25 m), các loài hiện diện phong phú hơn, đặc biệt là Pteria penguin và Charybdis japonica. Tại chân rạn, Colochirus quadrangularis xuất hiện với mật độ cao, khẳng định vai trò loài có thể là chỉ thị sinh thái trong điều kiện nền đáy ổn định ít bị tác động. Kết quả này phù hợp với các nghiên cứu tại Bạch Long Vĩ và Cát Bà, trong đó các loài như Colochirus quadrangularis thường phân bố ở vùng giữa và chân rạn trên nền đáy ổn định, phản ánh chất lượng môi trường đáy tốt (Thành, 2021), (Ngọc, 2020). Tuy nhiên, các loài có giá trị kinh tế như Atrina vexillum và Pteria peasei đang chịu áp lực khai thác gia tăng, nhất là tại tầng nông, làm gia tăng nguy cơ suy giảm nguồn lợi và mất cân bằng sinh thái.
4. KẾT LUẬN
Kết quả nghiên cứu về đặc điểm thành phần loài và phân bố sinh vật đáy là lần đầu tiên được công bố ở gò đồi ngầm Cô Tô - Thanh Lân. Kết quả đã ghi nhận 95 loài động vật đáy thuộc ba ngành chính: Mollusca, Arthropoda và Echinodermata, với sự khác biệt rõ rệt trong phân bố loài theo độ sâu và nền đáy như rạn san hô (12 - 15 m); đá tảng và cụm san hô (18 - 22 m); đáy cát bùn (25 - 35 m). Kết quả có chỉ ra mối liên quan giữa dạng sinh thái và nền đáy với một số nhóm loài đặc trưng như loài Mauritia arabica tập trung ở vùng rạn san hô nông (14 - 20 m), trong khi Atrina vexillum, Pteria penguin và Colochirus quadrangularis hiện diện chủ yếu ở tầng sâu (>25 m), nơi nền đáy cát bùn ổn định và giàu dinh dưỡng hữu cơ. Nghiên cứu cũng ghi nhận một số loài có giá trị kinh tế như Atrina vexillum, Pteria peasei và Mauritia arabica, là những đối tượng có tiềm năng quy hoạch bảo vệ và khai thác bền vững. Tuy nhiên, cần thận trọng với áp lực từ hoạt động khai thác không kiểm soát, đặc biệt tại các tầng nông dễ tiếp cận. Đây là cơ sở khoa học quan trọng để đề xuất các giải pháp bảo tồn HST đáy đặc thù theo tầng sâu và nền đáy, nhằm bảo vệ các loài đặc hữu - kinh tế và hướng tới phát triển kinh tế biển bền vững tại khu vực gò đồi ngầm Cô Tô - Thanh Lân.
Lời cảm ơn: Bài báo được hoàn thành dựa trên kết quả nghiên cứu của nhiệm vụ khoa học cấp Bộ Nông nghiệp và Môi trường “Điều tra tổng thể ĐDSH và nguồn lợi hải sản tại các bãi cạn, gò đồi ngầm ở vùng biển Việt Nam phục vụ công tác bảo tồn và phát triển bền vững ngành thuỷ sản Việt Nam” - thực hiện từ năm 2023 - 2025, do Viện Nghiên cứu Hải sản chủ trì thực hiện. Xin trân trọng cảm ơn Bộ NN&MT và Viện Nghiên cứu Hải sản đã tạo điều kiện để hoàn thành các nội dung nghiên cứu của bài báo.
Bùi Minh Tuấn1,*, Nguyễn Khắc Bát1, Hoàng Đình Chiều1, Lưu Xuân Hòa1, Phạm Trần Đình Nho1, Dư Văn Toán2
1Viện Nghiên cứu Hải sản, Bộ Nông nghiệp và Môi trường
2Viện Khoa học môi trường, biển và hải đảo, Bộ Nông nghiệp và Môi trường
(Nguồn: Bài đăng trên Tạp chí Môi trường, số 5/2025)
Tài liệu tham khảo
1. Bát, N. K. (2022). Mối quan hệ giữa cấu trúc nền đáy và mật độ sinh vật đáy vùng biển Vân Đồn - Quảng Ninh. Tạp chí Khoa học và Công nghệ Thủy sản, 18(3), 88 - 95.
2. Carpenter, K. E., & Niem, V. H. (Eds.). (1998a). FAO species identification guide for fishery purposes. The living marine resources of the Western Central Pacific. Volume 1: Seaweeds, corals, bivalves and gastropods, 686.
3. Carpenter, K. E., & Niem, V. H. (Eds.). (1998b). FAO species identification guide for fishery purposes. The living marine resources of the Western Central Pacific. Volume 2: Cephalopods, crustaceans, holothurians and sharks, 687-1396 p.
4. English, S., Wilkinson, C., & Baker, V. . (1997). Survey manual for tropical marine resources Australian Institute of Marine Science, 2nd ed, 390 pp.
5. Harris, P. T., Macmillan-Lawler, M., Rupp, J., & Baker, E. K. . (2014). Geomorphology of the oceans. Marine Geology, 352, 4–24.
6. Kent E. Carpenter, V. H. N. (1998). Species Identification Guide for Fishery Purposes: The Living Marine Resources of the Western Central Pacific (Vol. 2). FAO.
7. Khương, e. a. Đ. V. (2005). Nghiên cứu bổ sung cơ sở khoa học cho việc quy hoạch, quản lý các khu bảo tồn biển Cát Bà và Cô Tô. Báo cáo tổng kết. Viện Nghiên cứu Hải sản, 162.
8. Ngọc, T. T. B. (2020). Đặc điểm đa dạng thành phần loài động vật đáy ở vùng biển Cô Tô mở rộng. Tạp chí Khoa học Biển, 38(2), 45–58.
9. Nguyễn Văn Khương, P. M. T., & Nguyễn Thị Hòa (2015). Đặc điểm biến động mùa của sinh vật đáy tại vùng ven biển Quảng Ninh. Tạp chí Sinh học, 37 (2), 89 - 97.
10. Nguyễn Văn Quân, L. T. T. H., & Trần Văn Hùng (2012). Nghiên cứu ĐDSH sinh vật đáy tại khu vực Cô Tô – Thanh Lân, Quảng Ninh. Tạp chí Khoa học Biển và Hải đảo, 12(3), 45–54.
11. Okutani, T. E. (2000). Marine mollusks in Japan. Tokai University Press, 1173.
12. Pitcher, C. R., Ellis, N., Jebri, F., McLeod, I. M., Browne, M., & Dichmont, C. M. . (2016). Environmental surrogates to predict the distribution of biodiversity in the deep sea. Ecological Indicators, 61, 678–692.
13. Roberts, C. M., Bohnsack, J. A., Gell, F., Hawkins, J. P., & Goodridge, R. . (2002). Effects of marine reserves on adjacent fisheries. Science, 294(5548), 1920–1923.
14. Rogers, A. D. (2019). Chapter 23: Threats to Seamount Ecosystems and Their Management. In World Seas: An Environmental Evaluation. (Vol. III, pp. 427 - 451). Elsevier Ltd. https://doi.org/https://doi.org/10.1016/B978-0-12-805052-1.00018-8
15. Terrence, M., Gosliner, T. M., & Behrens, D. W. . (1996). Nudibranchs of the World. Monterey: Sea Challengers, 240.
16. Tuấn, L. M. (2019). Đánh giá ĐDSH sinh vật đáy theo độ sâu tại vùng biển Hòn Dáu – Hải Phòng. Tạp chí Khoa học Biển, 35(2), 54–62.
17. Thành, N. V. (2021). Đặc điểm phân bố sinh vật đáy theo nền đáy vùng biển ven bờ Đông Bắc Việt Nam. Tạp chí Sinh học và Biển, 39(1), 22–30.
18. Wessel, P., Sandwell, D. T., & Kim, S.-S. . (2010). The global seamount census. Oceanography, 23(1), 24 - 33.
Phụ lục. Danh mục thành phần loài động vật đáy tại khu vực gò đồi ngầm Cô Tô - Thanh Lân
|
TT |
Taxon |
Thời điểm ghi nhận |
Tổng số |
|
|
2023 |
2024 |
|||
|
A. |
Arthropoda |
5 |
16 |
21 |
|
I |
Malacostraca |
|
|
|
|
1 |
Decapoda |
|
|
|
|
|
Alpheidae |
|
|
|
|
1 |
Alpheus malabaricus (Fabricius, 1775) |
X |
|
X |
|
2 |
Athanas japonicus Kubo, 1936 |
|
X |
X |
|
|
Diogenidae |
|
|
|
|
3 |
Dardanus lagopodes (Forsskål, 1775) |
X |
|
X |
|
4 |
Diogenes dubius (Herbst, 1804) |
|
X |
X |
|
5 |
Paguristes ortmanni Miyake, 1978 |
|
X |
X |
|
|
Portunidae |
|
|
|
|
6 |
Charybdis (Charybdis) annulata (Fabricius, 1798) |
|
X |
X |
|
7 |
Charybdis japonica de Haan, 1835 |
|
X |
X |
|
8 |
Charybdis sp. |
|
X |
X |
|
9 |
Portunus (Monomia) argentatus (A.Milne-Edwards,1861) |
X |
|
X |
|
10 |
Portunus (Xiphonectes) hastatoides (Fabricius, 1798) |
X |
|
X |
|
11 |
Portunus pelagicus (Linnaeus, 1758) |
|
X |
X |
|
12 |
Portunus sanguinolentus (Herbst, 1783) |
|
X |
X |
|
13 |
Thalamita crenata (Rüppell, 1830) |
|
|
|
|
|
Stenopodidae |
|
|
|
|
14 |
Stenopus pyrsonotus Goy and Devaney, 1980 |
X |
|
X |
|
|
Hippolytidae |
|
|
|
|
15 |
Hippolyte ventricosa H.Milne Edwards, 1837 |
|
X |
X |
|
|
Panopeidae |
|
|
|
|
16 |
Eurypanopeus abbreviatus (Stimpson, 1860) |
|
X |
X |
|
|
Penaeidae |
|
|
|
|
17 |
Penaeus monodon Fabricius, 1798 |
|
X |
X |
|
|
Trapeziidae |
|
|
|
|
18 |
Trapezionida sp. |
|
X |
X |
|
2 |
Pleocyemata |
|
|
|
|
|
Epialtidae |
|
|
|
|
19 |
Chorilia sp. |
|
X |
X |
|
3 |
Stomatopoda |
|
|
|
|
|
Squillidae |
|
|
|
|
20 |
Oratosquilla oratoria (De Haan, 1844) |
|
X |
X |
|
II |
Maxillopoda |
|
|
|
|
4 |
Sessilia |
|
|
|
|
|
Chthamalidae |
|
|
|
|
21 |
Chthamalus stellatus (Poli, 1791) |
|
X |
X |
|
III |
Thecostraca |
|
|
|
|
5 |
Balanomorpha |
|
|
|
|
|
Balanidae |
|
|
|
|
22 |
Balanus calidus Pilsbry, 1916 |
|
X |
X |
|
B. |
Echinodermata |
7 |
13 |
19 |
|
IV |
Asteroidea |
|
|
|
|
6 |
Valvatida |
|
|
|
|
|
Goniasteridae |
|
|
|
|
23 |
Goniodiscaster scaber (Moebius, 1859) |
X |
|
X |
|
|
Oreasteridae |
|
|
|
|
24 |
Pentaceraster cumingi (Gray, 1840) |
|
X |
X |
|
V |
Crinoidea |
|
|
|
|
7 |
Comatulida |
|
|
|
|
|
Colobometridae |
|
|
|
|
25 |
Oligometra serripinna (Carpenter, 1881) |
|
X |
X |
|
|
Himerometridae |
|
|
|
|
26 |
Himerometra sp. |
X |
|
X |
|
|
Mariametridae |
|
|
|
|
27 |
Lamprometra palmata (Müller, 1841) |
|
X |
X |
|
28 |
Stephanometra indica (Smith, 1876) |
|
X |
X |
|
29 |
Stephanometra sp. |
|
X |
X |
|
VI |
Echinoidea |
|
|
|
|
8 |
Camarodonta |
|
|
|
|
|
Temnopleuridae |
|
|
|
|
30 |
Temnopleurus hardwickii (Gray, 1855) |
|
X |
X |
|
31 |
Temnopleurus reevesii (Gray, 1855) |
|
X |
X |
|
32 |
Temnopleurus toreumaticus (Leske 1778) |
X |
|
X |
|
33 |
Salmacis sphaeroides (Linnaeus, 1758) |
|
X |
X |
|
9 |
Diadematoida |
|
|
|
|
|
Diadematidae |
|
|
|
|
34 |
Diadema setosum (Leske, 1778) |
X |
X |
X |
|
VII |
Holothuroidea |
|
|
|
|
10 |
Dendrochirotida |
|
|
|
|
|
Cucumariidae |
|
|
|
|
35 |
Cercodemas anceps Selenka, 1867 |
X |
|
X |
|
36 |
Colochirus quadrangularis Troschel, 1846 |
X |
|
X |
|
37 |
Pentacta anceps (Selenka, 1867) |
|
X |
X |
|
|
Phyllophoridae |
|
|
|
|
38 |
Pentacta quadrangularis (Troschel, 1846) |
|
X |
X |
|
11 |
Holothuriida |
|
|
|
|
|
Holothuriidae |
|
|
|
|
39 |
Holothuria leucospilota (Brandt, 1835) |
|
X |
X |
|
VIII |
Ophiuroidea |
|
|
|
|
12 |
Ophiacanthida |
|
|
|
|
|
Ophiocomidae |
|
|
|
|
40 |
Ophiocoma erinaceus Müller & Troschel, 1842 |
X |
|
X |
|
C. |
Mollusca |
21 |
45 |
55 |
|
IX |
Bivalvia |
|
|
|
|
13 |
Adapedonta |
|
|
|
|
|
Solenidae |
|
|
|
|
41 |
Solen grandis Dunker, 1862 |
|
X |
X |
|
14 |
Arcida |
|
|
|
|
|
Arcidae |
|
|
|
|
42 |
Barbatia foliata (Forsskål, 1775) |
X |
X |
X |
|
43 |
Barbatia lima (Reeve, 1844) |
|
X |
X |
|
15 |
Cardiida |
|
|
|
|
|
Cardiidae |
|
|
|
|
44 |
Vasticardium orbita (Broderip & G. B. Sowerby I, 1833) |
|
X |
X |
|
16 |
Mytilida |
|
|
|
|
|
Modiolidae |
|
|
|
|
45 |
Modiolus philippinarum Hanley, 1843 |
|
X |
X |
|
|
Mytilidae |
|
|
|
|
46 |
Perna viridis (Linnaeus, 1758) |
X |
X |
X |
|
17 |
Ostreida |
|
|
|
|
|
Gryphaeidae |
|
|
|
|
47 |
Hyotissa hyotis (Linnaeus, 1758) |
|
X |
X |
|
48 |
Hyotissa inermis (G. B. Sowerby II, 1871) |
|
X |
X |
|
49 |
Hyotissa sp. |
|
X |
X |
|
|
Isognomonidae |
|
|
|
|
50 |
Isognomon isognomum (Linnaeus, 1758) |
|
X |
X |
|
51 |
Isognomon sp. |
X |
|
X |
|
|
Margaritidae |
|
|
|
|
52 |
Pinctada fucata (A. Gould, 1850) |
|
X |
X |
|
53 |
Pinctada imbricata Röding, 1798 |
|
X |
X |
|
54 |
Pinctada margaritifera (Linnaeus, 1758) |
X |
|
X |
|
55 |
Pinctada sp. |
|
X |
X |
|
|
Ostreidae |
|
|
|
|
56 |
Crassostrea sp. |
X |
X |
X |
|
57 |
Magallana gigas (Thunberg, 1793) |
X |
|
X |
|
58 |
Magallana nippona (Seki, 1934) |
X |
|
X |
|
59 |
Magallana sp. |
X |
X |
X |
|
|
Pinnidae |
|
|
|
|
60 |
Atrina vexillum (Born, 1778) |
X |
|
X |
|
18 |
Pectinida |
|
|
|
|
|
Anomiidae |
|
|
|
|
61 |
Anomiidae sp. |
|
X |
X |
|
|
Pectinidae |
|
|
|
|
62 |
Mimachlamys varia (Linnaeus, 1758) |
|
X |
X |
|
|
Spondylidae |
|
|
|
|
63 |
Spondylus sp. |
|
X |
X |
|
19 |
Pinnida |
|
|
|
|
|
Pinnidae |
|
|
|
|
64 |
Atrina pectinata (Linnaeus, 1767) |
|
X |
X |
|
20 |
Pteriida |
|
|
|
|
|
Pinnidae |
|
|
|
|
65 |
Atrina vexillum (Born, 1778) |
|
X |
X |
|
66 |
Pinna bicolor Gmelin, 1791 |
X |
|
X |
|
67 |
Pinna electrina (Linnaeus, 1758) |
|
X |
X |
|
|
Pteriidae |
|
|
|
|
68 |
Pteria brevialata (Dunker, 1872) |
|
X |
X |
|
69 |
Pteria peasei (Dunker, 1872) |
X |
X |
X |
|
70 |
Pteria penguin (Röding, 1798) |
X |
X |
X |
|
21 |
Venerida |
|
|
|
|
|
Chamidae |
|
|
|
|
71 |
Chama sp. |
|
X |
X |
|
|
Solenidae |
|
|
|
|
72 |
Trisidos kiyonoi (Makiyama, 1931) |
|
X |
X |
|
|
Veneridae |
|
|
|
|
73 |
Paratapes undulatus (Born, 1778) |
X |
|
X |
|
74 |
Periglypta chemnitzi (Hanley, 1844) |
X |
|
X |
|
X |
Gastropoda |
|
|
|
|
22 |
Aplysiida |
|
|
|
|
|
Aplysiidae |
|
|
|
|
75 |
Aplysiidae sp. |
|
X |
X |
|
23 |
Littorinimorpha |
|
|
|
|
|
Cerithiidae |
|
|
|
|
76 |
Cerithium sp. |
|
X |
X |
|
|
Cypraeidae |
|
|
|
|
77 |
Mauritia arabica asiatica Schilder & Schilder, 1939 |
X |
X |
X |
|
78 |
Monoplex mundus (Gould, 1849) |
X |
X |
X |
|
|
Ovulidae |
|
|
|
|
79 |
Crenavolva traillii (A. Adams, 1856) |
|
X |
X |
|
24 |
Neogastropoda |
|
|
|
|
|
Conidae |
|
|
|
|
80 |
Conus textile Linnaeus, 1758 |
X |
|
X |
|
|
Muricidae |
|
|
|
|
81 |
Chicoreus brunneus (Link, 1807) |
X |
X |
X |
|
82 |
Chicoreus ryukyuensis (Shikama, 1978) |
|
X |
X |
|
83 |
Ergalatax sp. |
|
X |
X |
|
84 |
Indothais sacellum (Gmelin, 1791) |
|
X |
X |
|
85 |
Mancinella echinata (Blainville, 1832) |
X |
X |
X |
|
86 |
Rapana rapiformis (Born, 1778) |
X |
|
X |
|
87 |
Reishia clavigera (Küster, 1860) |
|
X |
X |
|
88 |
Reishia jubilaea (K. S. Tan & Sigurdsson, 1990) |
|
X |
X |
|
89 |
Thais sp. |
|
X |
X |
|
90 |
Urosalpinx cinerea (Say, 1822) |
|
X |
X |
|
|
Nassariidae |
|
|
|
|
91 |
Nassarius exilis (Powys, 1835) |
|
X |
X |
|
25 |
Nudibranchia |
|
|
|
|
|
Chromodorididae |
|
|
|
|
92 |
Chromodoris lineolata (van Hasselt, 1824) |
|
X |
X |
|
93 |
Doriprismatica atromarginata (Cuvier, 1804) |
X |
X |
X |
|
|
Polyceridae |
|
|
|
|
94 |
Roboastra sp. |
|
X |
X |
|
26 |
Vetigastropoda |
|
|
|
|
|
Tegulidae |
|
|
|
|
95 |
Tectus pyramis (Born, 1778) |
|
X |
X |
|
|
Tổng số loài |
33 |
74 |
95 |